![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
38 Cards in this Set
- Front
- Back
|
8.6.2 De basale ganglia
Zoals het cerebellum hebben de basale ganglia geen directe efferente connecties met het ruggenmerg en spelen ze een indirecte rol in de bewegingscontrole.
In tegenstelling tot het cerebellum krijgen de basale ganglia |
geen directe sensorische input vanuit het ruggenmerg en zijn ze daarvoor afhankelijk van de cerebrale cortex.
Hun output gaat via de thalamus naar de prefrontale, premotorische en motorische cortex (Ghez & Gordon, 1995b).
|
|
|
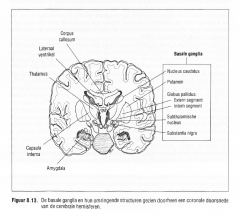
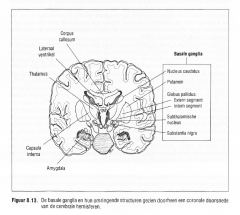
De basale ganglia bestaan uit vijf subcorticale grijze kernen, gelegen rond de thalamus en de hypothalamus , en zijn met elkaar verbonden door talrijke interconnecties.
In de basale ganglia onderscheiden we
n p g s s |
-de nucleus caudatus, -het putamen, -de globus pallidus, -de subthalamische kern en -de substantia nigra
(Figuur 8.13.).
|
|
|
Figuur 8.13.
De basale ganglia en hun omringende structuren gezien doorheen een coronale doorsnede
|
|
|
|
-de nucleus caudatus, -het putamen, -de globus pallidus, -de subthalamische kern en -de substantia nigra
De eerste drie behoren tot |
het telen- en diencephalon, terwijl de laatste twee tot het mesencephalon behoren (Rothwell, 1994).
|
|
|
De nucleus caudatus en het putamen zijn filogenetisch het |
jongst en ontwikkelen uit dezelfde telencephale structuur (waaruit ook de cerebrale cortex zich ontwikkelde).
Ze zijn opgebouwd uit dezelfde celtypes en worden samen het neostriatum genoemd. |
|
|
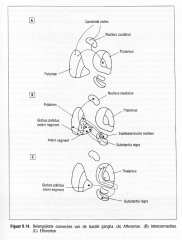
Figuur 8.14 .
Belangrijkste connecties van de basale ganglia.
(A) Afferenten . (B) lnterconnecties . (C) Efferenten.
|
|
|
|
De globus pallidus ontwikkelde zich uit het diencephalon en wordt ook wel paleostriatum genoemd.
De globus pallidus heeft een totaal andere cellulaire structuur als het neostriatum en bestaat uit |
een intern en extern egment.
|
|
|
De subthalamische nucleus ligt onder de thalamus.
De substantia nigra heeft twee zones : |
-een ventrale bleke zone, de pars reticulata, die cytologisch op de globus pallidus lijkt ; -en een dorsale donker gepigmenteerde zone, de pars compacta.
Hetzelfde geldt voor het putamen en de nucleus caudatus (Beatty, 1995 ; Bradshaw & Mattingley, 1995 ; Ghez & Gordon, 1995b).
|
|
|
Bijna alle afferente banen naar de basale ganglia eindigen in bet neostriatum.
De belangrijkste bronnen zijn de thalamus en de cerebrale cortex (Figuur 8.14.A.).
Van daaruit wordt naar de globus pallidus en de substantia nigra geprojecteerd die de belangrijkste outputkernen van de basale ganglia vormen (Figuur 8.14.B .).
Deze efferente banen eindigen in verschillende thalamuskernen die op hun beurt naar verschillende frontale cmtexareas projecteren (Figuur 8.14.C.). |
|
|
|
Zowel de externe connecties als de interconnecties tussen de kernen van de basale ganglia zijn topografisch georganiseerd, parallel aan de somatotopische organisatie van de overige motorische systemen.
Bepaalde cortexareas projecteren naar bepaalde gebieden van het neostriatum die op hun beurt |
naar bepaalde gebieden van de globus pallidus en de substantia nigra projecteren (Ghez & Gordon, 1995b). |
|
|
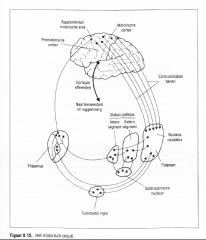
De talrijke connecties tussen de basale ganglia en de cerebrale cortex bestaan uit vijf functioneel en anatomisch onderscheidbare circuits die verantwoordelijk zijn voor de regulatie van bewegingen met de ledematen, oogbewegingen en meer complexe cognitieve gedragingen.
Het best begrepen circuit is het motorisch circuit (Figuur 8.15.) dat bestaat uit de corticostriatale baan die ontspringt ter hoogte van die cortexareas betrokken bij de bewegingscontrole (SMA, premotorische zone, primaire motorische cortex, somatosensorische cortex en de supeiieure pariëtale lob).
|
|
|
|
Het best begrepen circuit is het motorisch circuit (Figuur 8.15.) dat bestaat uit de corticostriatale baan die ontspringt ter hoogte van die cortexareas betrokken bij de bewegingscontrole (SMA, premotorische zone, primaire motorische cortex, somatosensorische cortex en de supeiieure pariëtale lob).
Deze eindigt in |
het putamen dat directe en indirecte connecties heeft met de globus pallidus en de substantia nigra waarvan de output via de thalamus naar de SMA en de premotorische zone projecteert.
|
|
|
Deze areas hebben reciproque interconnecties en connecties naar de primaire motorische cortex en deze zones hebben alle drie descenderende projecties naar de |
hersenstam en het ruggenmerg.
Via deze drie descenderende banen oefenen de basale ganglia dus invloed uit op de lichaamshouding en de bewegingen van de ledematen (Ghez
|
|
|
Het motorisch circuit tussen het putamen enerzijds en de globus pallidus en de substantia nigra anderzijds heeft twee routes, een |
indirecte en een directe (Hallett, 1993 ; Wichmann
De directe route is een faciliterend feedback circuit voor het volhouden of faciliteren van een beweging.
De indirecte route synapteert in de subthalarnische nucleus en werkt inhiberend zodat ongewenste motorische programma's onderdrukt kunnen worden.
|
|
|
De directe route is een faciliterend feedback circuit voor het volhouden of faciliteren van een beweging.
De indirecte route synapteert in de subthalarnische nucleus en werkt inhiberend zodat ongewenste motorische programma's onderdrukt kunnen worden.
De basale ganglia kunnen dus |
specifieke motorische synergieën selecteren (via de directe route) of inhiberen (via de indirecte route).
|
|
|
Wanneer willekeurige bewegingen gefaciliteerd worden, zullen de spinale reflexen geïnhibeerd worden.
Een overactieve indirecte of een onderactieve directe route (zoals bij de ziekte van Parkinson) zal |
de corticale activiteit onderdrukken en het starten van bewegingen bemoeilijken, maar tegelijk de responsiviteit van de spinale reflexen verhogen, met de Parkinsoniaanse akinesie en rigiditeit (door hypertonie) tot gevolg .
Een onderactieve indirecte en een overactieve directe route zal door stimulatie van de cortex leiden tot een onvrijwillige overvloed aan willekeurige bewegingen, zoals bij de ziekte van Huntington, hoewel bij deze aandoening de Parkinoniaanse akinesie en bradykinesie ook voorkomen (Bradshaw & Mattingley , 1995). |
|
|
Een onderactieve indirecte en een overactieve directe route zal door stimulatie van de cortex leiden tot |
een onvrijwillige overvloed aan willekeurige bewegingen, zoals bij de ziekte van Huntington, hoewel bij deze aandoening de Parkinoniaanse akinesie en bradykinesie ook voorkomen (Bradshaw & Mattingley , 1995). |
|
|
De basale ganglia kunnen als wat beschouwd worden?
|
Als een soort automatische piloot beschouwd worden (Steg & Johnels, 1993), die op een onbewust niveau de verschillende bewegingsprogramma's coördineert en zo de cortex (de piloot die bewust willekeurige beslissingen neemt) van deze taak ontslaat.
De basale ganglia opereren dan ook meer in de context van goedgekende semi-automatische bewegingsprogramma 's die weinig of geen aandacht vragen, en minder in een context waarin een nieuwe, ongekende of moeilijke taak een constante monitoring en aandacht vereist. |
|
|
De basale ganglia opereren dan ook meer in de context van |
goedgekende semi-automatische bewegingsprogramma 's die weinig of geen aandacht vragen, en minder in een context waarin een nieuwe, ongekende of moeilijke taak een constante monitoring en aandacht vereist. |
|
|
Voor het uitvoeren van dergelijke geoefende bewegingssequensen werken de basale ganglia samen met |
de SMA en verzekeren daarin een vloeiende overgang tussen opeen volgende bewegingsonderdelen.
De SMA genereert een premotorische tonische neuronale activiteit (Bereitschaftspotential) die bij het starten van de beweging onmiddellijk ophoudt. |
|
|
De basale ganglia genereren een |
fasische activiteit tijdens bewegingen en naar het einde toe. |
|
|
Deze fasische activiteit fungeert als |
een intern motorisch signaal dat de tonische premotorische activiteit van de SMA stopzet zodat het bewegingsonderdeel uitgevoerd kan worden, en dat het startsein geeft voor de premotorische activiteit voor het daaropvolgende bewegingsonderdeel.
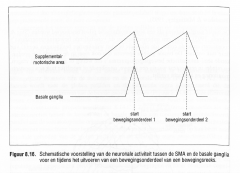
De uitvoering van de beweging lokt bij de basale ganglia een volgend fasisch signaal uit dat op zijn beurt de premotorische activiteit voor het daaropvolgende bewegingsonderdeel uitdooft zodat dit uitgevoerd kan worden enzovoort, totdat de volledige bewegingsreeks uitgevoerd is (Figuur 8.16.). |
|
|
Schematische voorstelling van de neuronale activiteit tussen de SMA en de basale ganglia voor en tijdens het uitvoeren van een bewegingsonderdeel van een bewegingsreeks.
|
|
|
|
De basale ganglia reguleren dus de continue activiteit van de SMA door deze |
op het gepaste ogenblik uit en aan te schakelen bij iedere nieuwe fase van een bewegingsprogramma (Brotchie, Iansek & Home, 1991ab ; Georgiou et al" 1993).
|
|
|
Dit kan een verklaring vormen van de problemen die parkinsonpatiënten hebben met het starten en stoppen van bewegingen.
Het belang van deze intern gegenereerde cues verdwijnt bij onbekende nieuwe bewegingen waarvoor veel aandacht vereist is (Bradshaw & Mattingley, 1995).
Terwijl de basale ganglia vooral betrokken zijn bij zelf-geïnduceerde bewegingen, lijken ze daarentegen niet van belang bij |
de initiatie van bewegingen die door een externe stimulus worden uitgelokt.
Vandaar dat parkinsonpatiënten problemen hebben met het starten van bewegingen die niet door een externe stimulus worden uitgelokt (Ghez & Gordon, 1995b). |
|
|
Aandoeningen van de basale ganglia produceren tal van bewegingsstoornissen:
|
-De ziekte van Parkinson is de meest voorkomende aandoening van de basale ganglia en werd genoemd naar de Engelsman die het voor het eerst beschreef in 1817.
-Ook de ziekte van Huntington , een erfelijke aandoening die gekenmerkt wordt door het optreden van een groot aantal onwillekeurige, snelle, gecompliceerde, schokkende bewegingen ( chorea), is het gevolg van een stoornis in het neurotransmittersysteem van de basale ganglia |
|
|
De ziekte van Parkinson
De ziekte van Parkinson is de meest voorkomende aandoening van de basale ganglia en werd genoemd naar de Engelsman die het voor het eerst beschreef in 1817.
Deze progressieve ziekte treft vooral patiënten op late middelbare leeftijd en wordt gekenmerkt door negatieve en positieve symptomen.
De voornaamste negatieve symptomen zijn : |
-verlies aan spontane bewegingen ( akinesie) ; vertraagd initiëren van bewegingen, vooral wanneer ze niet door een externe stimulus worden uitgelokt met als gevolg een verlaagde reactiesnelheid ( hypokinesie ) ; -vertraagd uitvoeren van bewegingen (bradykinesie ).
Andere negatieve symptomen zijn een zwakke monotone dysartrische spraak met neiging tot propulsie (plotselinge versnelling), micrografie (een progressieve afname in grootte en snelheid van het schrift over enkele lijnen) en een vertraagde gang gekenmerkt door schuifelende kleine pasjes.
|
|
|
De ziekte van Parkinson.
De belangrijkste positieve symptomen zijn |
rigiditeit en tremo.
Door de verhoogde tonus worden de spieren rigide, wat zich manifesteert door een weerstand tegen pas ieve beweging in de extensoren en vooral in de flexoren, met houdingsproblemen tot gevolg.
Deze weerstand is van het soepele 'loden pijp' -type, hoewel ze in interactie met tremor een soort 'tandrad '-kwaliteit kan hebben. |
|
|
De rigiditeit bij de ziekte van Parkinson is ander dan de rigiditeit bij pasticiteit (na letsels van de motorische cortex en/of de cortison spinale baan) die gepaard gaat met |
het zogenaamde 'knipmes' -fenomeen.
Subtiele rigiditeit kan uitgelokt worden door bewegingen met het contralaterale lichaamsdeel. |
|
|
De tremor (vooral in handen en vingers) komt voor bij de meerderheid van de parkinsonpatiënten, maar zeker niet bij allemaal (Marsden. 1994).
Deze tremor treedt enkel op in rust (rusttremor) en verdwijnt bij het gebruik van het lichaamsdeel of tijdens de slaap, dit in tegenstelling tot
|
de zogenaamde intentietremor bij cerebellaire letsels die maximaal is tijden willekeurige doelgerichte bewegingen.
|
|
|
De rusttremor kan toenemen in één arm bij het gebruik van de contralaterale arm of in stresssituaties (bijvoorbeeld bij hoofdrekenen).
De tremor kan onderdrukt worden door de wil, tijdens de slaap of tijdens volledige relaxatie.
Dit symptoom is het minst gevoelig voor |
antiparkinsoniaanse medicatie en correleert in tegenstelling tot bradykinesie weinig met het cognitief functioneren.
Wanneer ze niet behandeld worden, nemen de symptomen gradueel toe, totdat patiënten uiteindelijk niet meer vrijwillig ku nnen bewegen (Bradshaw & Mattingley, 1995, Ghez |
|
|
In de jaren vijftig was er een doorbraak in de kennis over de biochemische basis van deze ziekte met de ontdekking dat er een groot tekort aan dopamine is in de hersenen van parkinsonpatiënten.
Dit tekort is het gevolg van |
de degeneratie van dopaminerge cellen in de substantia nigra die naar het neostriatum projecteren, waar ze cruciaal zijn voor de informatieverwerking functie van de basale ganglia.
Toediening van L-dopa (een precursor van dopamine) verhoogt de synthese van dopamine in de neuronen van de substantia nigra (die nog niet allemaal gedegenereerd zijn) en heeft een uitgesproken therapeutisch effect.
|
|
|
Deze behandeling geneest de ziekte echter niet want het effect van L-dopa neemt af met de tijd vermoedelijk door |
een progressieve degeneratie van de resterende dopaminerge cellen in de sub tantia nigra (Ghez & Gordon, 1995b).
|
|
|
De ziekte van Huntington.
Ook de ziekte van Huntington , een erfelijke aandoening die gekenmerkt wordt door het optreden van een groot aantal onwillekeurige, snelle, gecompliceerde, schokkende bewegingen ( chorea), is het gevolg van |
een stoornis in het neurotransmittersysteem van de basale ganglia.
Een verlies van kleine cholinerge neuronen en GABA bevattende neuronen zou tot een desinhibitie leiden van de dopaminerge cellen van de substantia nigra, wat op zijn beurt een inhibitie tot gevolg heeft van de projecties vanuit de globus pallidus naar de thalamus.
|
|
|
Een verlies van kleine cholinerge neuronen en GABA bevattende neuronen zou tot een desinhibitie leiden van de dopaminerge cellen van de substantia nigra, wat op zijn beurt |
een inhibitie tot gevolg heeft van de projecties vanuit de globus pallidus naar de thalamus.
Bij de ziekte van Huntington is er dus een tekort aan acetylcholine en GABA en een teveel aan dopamine in de ba ale ganglia.
|
|
|
Bij de ziekte van Huntington is er dus |
een tekort aan acetylcholine en GABA en een teveel aan dopamine in de ba ale ganglia.
Toediening van L-dopa leidt door een verder verstoren van het evenwicht tot een toename van de symptomen.
Anderzijds kunnen parkinsonpatiënten ook Huntington-achtige symptomen ontwikkelen door een te hoge toediening van L-dopa (Beatty, 1995).
Een correct evenwicht tussen de verschiJlende neurotransmittersystemen van de basale ganglia lijkt bijgevolg crnciaal te zijn voor een normaal motorisch functioneren (Beatty, 1995). |
|
|
Toediening van L-dopa leidt door een verder verstoren van het evenwicht tot een toename van de symptomen.
Anderzijds kunnen parkinsonpatiënten ook |
Huntington-achtige symptomen ontwikkelen door een te hoge toediening van L-dopa (Beatty, 1995).
Een correct evenwicht tussen de verschiJlende neurotransmittersystemen van de basale ganglia lijkt bijgevolg cruciaal te zijn voor een normaal motorisch functioneren (Beatty, 1995). |
|
|
Een correct evenwicht tussen de verschiJlende neurotransmittersystemen van de basale ganglia lijkt |
bijgevolg cruciaal te zijn voor een normaal motorisch functioneren (Beatty, 1995). |