![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
45 Cards in this Set
- Front
- Back
|
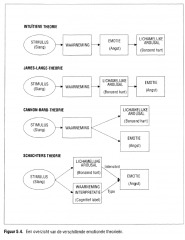
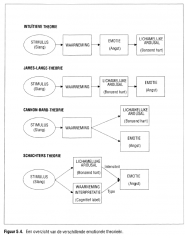
5.4 Theorieën over emotie
|
een omgekeerd verband .
|
|
|
Charles Darwin (1872) observeerde dat de lichamelijke expressie van een emotie onze gevoelens kunnen versterken : "He who gives violent gesture increases his rage."
Darwin beklemtoonde vooral de zelfwaarneming van de aangezichtsexpressie als |
het ontstekingsmechanisme van de emotionele ervaring. |
|
|
William James (1890) argumenteerde dat emotie niets meer of niets minder is dan |
de waarneming van perifere fysiologische veranderingen en schreef : "We feel sorry because we cry, afraid because we tremble." |
|
|
James stelde dat een emotie optreedt dankzij |
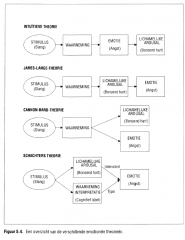
de waarneming van de lichamelijke reacties die optreden naar aanleiding van een emotionele stimulus (Figuur 5.4.).
|
|
|
Figuur 5.4. Een overzicht van de verschillende emotionele theorieën. |
|
|
|
James meende ook dat een specifieke emotionele ervaring af te leiden was uit een specifieke constellatie van lichamelijke veranderingen.
Het gevoel van angst is in feite |
het gevoel van een snelkloppend hart, oppervlakkige en snelle ademhaling, 'kippenvel' en bevende ledematen.
Woede is het gevoel van een pompende borst, rood aangelopen gezicht, opengesperde neusgaten en op elkaar geklem de kaken.
|
|
|
Een argument voor deze hypothese wordt gezocht in het feit dat mensen van over de gehele wereld hun emotie in termen van …….veranderingen beschrijven |
lichamelijke veranderingen beschrijven en dat er een vrij grote concensus is over het soort van veranderingen geassocieerd met elke emotie (Rime, Phillipot & Ci amolo, 1990).
Ook gelaatsuitdrukkingen die een
|
|
|
Onderzoek heeft tevens aangetoond dat het vormen van een emotionele gelaatsuitdrukking fysiologische reacties in de rest van het lichaam teweegbrengt die gelijken op de reacties die worden geproduceerd wanneer deze emotie effectief wordt gevoeld (Ekman, Levenson & Friesen, 1983 ; Leven on, 1992).
Het is de reactie van |
het sympathisch deel van ons autonoom zenuwstelsel dat de lichamelijke opwinding produceert , suggereerde James.
|
|
|
De sympathische reactie is een |
reflexmatige respons op de waarneming van emotie-uitlokkende stimuli.
|
|
|
Hoewel James ook cerebrale vormen van lust/onlust aanvaardde die geen (perceptie van) lichamelijke veranderingen noodzaakte, meende hij dat de emotie vooral wordt veroorzaakt door |
feedback van afferente zenuwvezels die de sympathische reacties van verschillende interne organen van thorax en abdomen (ook wel viscerale organen of kortweg viscera genoemd) en bloedvaten naar het centrale zenuwstelsel leiden. |
|
|
De Deense fysioloog Carl Lange publiceerde haast terzelfder tijd een gelijkaardige theorie ( 1885) waarbij hij de rol van veranderingen in constrictie en dilatatie van de bloed vaten benadrukte.
Daarom staat deze contra-intuïtieve theorie over emotie algemeen bekend als |
de James-Lange-theorie. |
|
|
De James-Lange-theorie.
Argumenten voor deze theorie vindt men bij patiënten met ruggenmerglaesies. Deze patiënten geven aan dat |
de intensiteit van hun emoties sterk is verminderd.
Dit geldt vooral bij patiënten met hoge laesies van het ruggenmerg die als gevolg hiervan een groot gedeelte van hun viscerale zintuiglijke waarnemingen moeten missen. De dwarslaesie patiënten beschrijven hun emoties als killer en zwakker dan voor het ongeval (Hohmann , 1966). |
|
|
Cannon meende dat de lichamelijke veranderingen niet grondig genoeg van elkaar verschillen om |
de verschillende emotionele ervaringen uit te lokken.
Een verhoogd hartritme kan toch evengoed voorkomen bij angst, woede als verliefdheid , en huilen kun je bij verdriet maar toch ook van geluk ? |
|
|
Dergelijke diffuse informatie vanuit het lichaam kon volgens Cannon de emotionele beleving niet verklaren en hij argumenteerde dat het scheiden van de viscera van het centraal zenuwstelsel (zodat feedback onmogelijk wordt) het hebben van emotionele gevoelens |
niet beïnvloedt.
|
|
|
Cannon verlamde het sympathisch zenuwstelsel in enkele katten door alle zenuwverbindingen tussen de sympathische neuronen en de interne organen en bloedvaten te verbreken.
Volgens de James-Lange-theorie zouden deze katten niet meer in staat mogen zijn emoties te ervaren aangezien hun sympathisch zenuwstelsel niet meer werkte.
Cannon stelde echter vast dat de dieren (in aanwezigheid van een blaffende hond) |
nog steeds duidelijke emotionele gedragingen vertoonden zoals blazen , grommen, platleggen van de oren, tonen van de tanden en het dreigen met de klauw, behalve het rechtzetten van de haren, een respons die normaal wordt veroorzaakt door de (afgesneden) sympathische ganglia. |
|
|
Aanhangers van de James-Lange-theorie opperden echter dat Cannon verkeerde conclusies trok.
Ze stelden dat de James-Lange-theorie verwees naar een 'gevoel' en niet naar de uiterlijke manifestatie van een emotie (er is uiteraard geen enkele manier om te weten te komen wat de katten werkelijk 'voelden').
Cannon argumenteerde dat het zeer onwaarschijnlijk was dat |
dergelijke gedragingen zich zouden manifesteren zonder dat de dieren zich ook werkelijk bedreigd zouden voelen.
|
|
|
Bovendien is het ook helemaal niet zo dat ruggenmerg laesies een emotioneel vacuüm creëren: |
ruggenmerglaesies kunnen de viscerale feedback nooit geheel onderbreken gezien de afferente parasympathische banen van de tiende craniale zenuw, de nervus vagus, die naar de hersenstam projecteren , intact blijven. |
|
|
5.4.2 Centrale theorieën
|
fysiologische arousal en de emotionele ervaring gebeuren terzelfdertijd (Figuur 5.4.).
De waarneming van de emotionele stimulus wordt simultaan zowel naar de hersenen (subjectieve ervaring van de emotie) al naar het sympathisch zenuwstelsel (lichamelijke arousal) gestuurd.
|
|
|
Figuur 5.4. Een overzicht van de verschillende emotionele theorieën. |
|
|
|
Cannon beklemtoonde het belang van een meer centrale, met name subcorticale rol in de mediatie van emotie.
Cannon steunde zich hiervoor op de bevindingen van zijn collega Philip Bard (1934) die vaststelde dat gedecorticeerde katten (met een intact diencephalon waaronder thalamus en hypothalamus) sterke emotionele reacties op schijnbaar triviale stimuli vertoonden : |
"sham rage".
Gedecerebreerde katten (waarbij naast het telencephalon ook het diencephalon van het mesencephalon wordt gescheiden) vertoonden deze reacties niet. Vooral de stimulatie van de hypothalamus kon het emotioneel gedrag uitlokken.
|
|
|
Deze studies leken erop te wijzen dat thalamus en hypothalamus de neurale circuits voor de uitdrukking van emotioneel gedrag bevatten, en volgens Cannon activeerden de waargenomen stimuli via de thalamus, de hypothalamus.
Gezien de hypothalamus het endocrien systeem en het autonoom zenuwstelsel controleert, kan de hypothalamus in vrijwel alle orgaansystemen veranderingen bewerkstelligen (emotionele uitdrukking).
Cannon meende ook dat een activatie van de cerebrale cortex door de hypothalamus |
de bewuste ervaring van de emotie veroorzaakte (emotionele ervaring), maar hij zag geen corticale rol in de (emotionele) interpretatie van de stimuli.
|
|
|
James Papez (1937) nam de centrale notie van de Cannon-Bard-theorie over, maar situeerde ze in een breder anatomisch netwerk : |
het limbisch systeem (zie 'Limbische structuren ' in het eerste hoofdstuk).
|
|
|
Paul Broca (1878) beschreef als eerste de limbische lob ('Je grand lobe limbique') als een set van verwante neurale structuren die gelegen zijn aan de mediale basis van de hersenen. Het woord limbisch is afgeleid van het Latijnse limbus, wat grens betekent.
Met deze grens wordt de scheiding bedoeld tussen de |
fylogenetisch oudere delen van de hersenen en de fylogenetisch jongere neocortex.
|
|
|
Volgens Broca bestond de limbische lob uit de gyrus hippocampus en parahippocampus, de gyrus cinguli en de gyrus subcallosalus.
Broca zag in het limbisch systeem vooral een |
anatomisch concept dat gezien de nabijheid van het olfactorisch systeem een olfactorische functie werd toegeschreven (rhinencephalon of reukhersenen). |
|
|
James Papez (1937) onderzocht de functie van de limbische structuren en benadrukte de rol van deze hersenregio als de neurale basis voor de emotie.
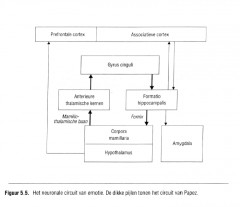
Papez stelde een ring van fylogenetisch primitieve cortex (de limbische lob van Broca) voor als een baansysteem |
(gyrus cinguli - formatio hippocampalis - corpora mamillaria - anterieure thalarnische kernen - gyrus cinguli)
waardoor de emotie tot het bewustzijn kan doordringen. |
|
|
Dit baansysteem wordt het circuit van Papez genoemd (Figuur 5.5.). |
|
|
|
Ook Papez zag de hypothalamus als de belangrijkste effector van de emotie die de ontvangen stimuli van een affectieve kleur voorzag.
Hij benadrukte echter de sterke invloed van |
de limbische structuren en be schouwde de gyrus cinguli als het corticale gebied verantwoordelijk voor de emotionele ervaring.
|
|
|
Papez stelde dat de neocortex de hypothalamus beïnvloedt via connecties van de gyrus cinguli met de hippocampus. De hippocampus verwerkt de informatie afkomstig van de corticale gyrus cinguli en projecteert die via de fomix (een belangrijke bundel efferente zenuwvezels die vertrekt vanuit de hippocampus) naar de (vlak achter de hypothalamus gelegen) |
corpora mamillaria.
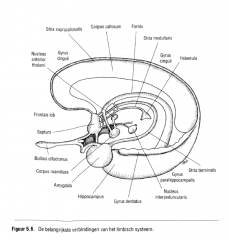
Op zijn beurt voorziet de hypothalamus in de informatie naar de gyrus cinguli via een baansysteem van de corpora mamillaria naar de anterieure thalamische kernen (de mamillothalarnische tractus) en van de anterieure thalamische kernen naar de gyrus cinguli (Figuur 5.6.). |
|
|
Figuur 5.6. De belangrijkste verbindingen van het limbisch systeem. |
|
|
|
In 1948 voegde Yakovlev de basolaterale componenten (orbitofrontale, insulaire en anterieure temporale cortices, de amygdala en de dorsomediale nucleus van de thalamus ) bij de basomediale structuren en Paul MacLean ( 1949) suggereerde dat |
de term limbisch systeem gebruikt kan worden om de verzameling van structuren te beschrijven die schijnbaar bijdragen tot de perceptie en expressie van emotie.
|
|
|
MacLean voegde ook het septum en delen van de basale ganglia toe aan het 'visceraal brein ' dat instond voor emotie en overleving.
Bij MacLean stond de hippocampus en niet de |
gyrus cinguli centraal als de corticale zetel van emotionele ervaring.
|
|
|
Deze korte historiek is illustratief voor het feit dat ook vandaag nog het concept 'limbisch systeem' verschillende betekenissen kan hebben voor verschillende disciplines.
Bovendien blijkt uit recenter onderzoek dat bepaalde structuren die aanvankelijk tot het limbisch systeem werden gerekend, andere dan emotionele functies hebben, zoals |
de hippocampus.
Andere limbische structuren zoals de amygdala blijken inmiddels veel belangrijker voor de opbouw van emotioneel gedrag. Gezien deze heterogene functionaliteit spreken we liever van limbische structuren dan van een limbisch systeem. |
|
|
Ondanks Cannons tegenwerpingen zijn er onvoldoende argumenten om de James-Lan ge-theorie en meer bepaald het belang van de viscerale feedback te verwerpen en blijft de feedbackidee (bewuste emotie komt na perifere fysiologische veranderingen) een belangrijke plaats innemen in de theorievorming rond emotie.
Een belangrijke nuance op de theorie van James en Lange werd voorgesteld door |
Stanley Schachter |
|
|
Hoewel Schachter, net zoals James en Lange, geloofde dat de emotionele ervaring groeit uit de waarneming van lichamelijke veranderingen, geloofde hij net als Cannon en Bard dat emotionele fysiologische reacties zeer sterk gelijkend zijn.
Schachter stelde dat een |
bewuste (corticale) interpretatie (cognitief label) nodig is om de arousal te kunnen verklaren.
Bij de mens speelt de cortex een belangrijke rol bij de analyse en interpretatie van stimuli die |
|
|
Farmacologisch geïnduceerde viscerale activatie produceert geen echte emotie, maar creëert 'alsof'-gevoelens die slechts betekenis krijgen als ze worden vergezeld door |
een 'cognitieve omgeving '. |
|
|
Schachter injecteerde zijn proefpersonen met epinephrine (een stof die het hartritme doet stijgen en ook andere lichamelijke effecten produceert die worden geassocieerd met hoge arousal) of met een placebo (inactieve substantie) en stelde hen vervolgens aan verschillende emotie-inducerende conditie bloot.
Hij stelde vast dat |
het effect van epinephrine alleen geen specifieke emotie produceerde (de subjecten voelden zich enkel wat 'nerveus').
|
|
|
Wanneer het effect van de epinephrine echter werd gecombineerd met een emotie-inducerende omgeving, bleek de intensiteit van de emotionele ervaring verhoogd.
Zoals door deze theorie werd voorspeld, bleek de emotie van de subjecten afhankelijk van |
de omgeving, maar werd de intensiteit ervan verhoogd door de epinephrine.
|
|
|
De met epinephrine geïnjecteerde proefpersonen manifesteerden en rapporteerden meer woede wanneer ze werden beledigd , meer angst tijdens het kijken naar een griezelige film en meer hilariteit bij het bekijken van een komedie dan de met placebo geïnjecteerde proefpersonen .
Dit emotieversterkend effect trad enkel op indien |
de proef personen niet op voorhand over de fysiologische effecten van de epinephrine werden geïnformeerd .
|
|
|
Een hoge fysiologische arousal zal een emotie slechts versterken indien de mensen geloven dat de arousal wordt veroorzaakt door |
de externe situatie.
Schachter besloot dat waarneming en analyse van de omgeving het type aangevoelde emotie bepalen en dat de sensorische feedback van de mate van lichamelijke arousal de aangevoelde intensiteit van de emotie beïnvloedt (Figuur 5.4.). |
|
|
Figuur 5.4. Een overzicht van de verschillende emotionele theorieën. |
|
|
|
Gebaseerd op deze studies nuanceerde Schachter de James-Lange-theorie naar een cognitieve arousal-theorie van emoties die stelt dat een emotionele ervaring gebaseerd is op |
specifieke cognitieve attributies ten aanzien van een eerder diffuse fysiologische arousal.
|
|
|
De cortex creëert een cognitieve respons op de perifere informatie die overeenkomt met de sociale context en de verwachtingen van het individu .
Een aangevoelde emotie wordt dus primair bepaald door de omgevingscontext waarin de arousal zich voordoet.
Analyse en interpretatie van een veelal complexe context impliceert uiteraard een belangrijke inbreng van |
hogere corticale areas.
De cortex dient over een feedforwardsysteem te beschikken om het autonoom zenuwstelsel en dus ook de viscera te controleren (viscerale efferenten) . |
|
|
Bovendien bestaan er neuronale banen die een feedbacksysteem vormen en de informatie vanuit het lichaam terng naar het brein voeren (viscerale afferenten).
De nervus vagus, de tiende craniale zenuw met een belangrijk parasympathisch -visceraal deel dat bestemd is voor de bezenuwing van de organen, is een goede kandidaat voor |
een dergelijk feedbacksysteem.
|
|
|
De afferenten van de nervus vagus synapteren in de hersenstam vooral ter hoogte van de kern van de tractus solitarius.
Deze nucleus solüarius projecteert naar de amygdala (en naar de hypothalamus) en ook naar de insula (Reili).
Amygdala en insula projecteren naar temporale, pariëtale en frontale corticale regio's.
Vermoed wordt dat dit vagaal-nucleus solitarius-amygdala-insulair baansysteem |
verantwoordelijk is voor de viscerale feedback (Heilman, Bowers & Valenstein, 1993). |
|
|
Het feedback systeem: |
-De afferenten van de nervus vagus synapteren in de hersenstam vooral ter hoogte van de kern van de tractus solitarius.
-Deze nucleus solitarius projecteert naar de amygdala (en naar de hypothalamus) en ook naar de insula (Reili).
-Amygdala en insula projecteren naar temporale, pariëtale en frontale corticale regio's.
Vermoed wordt dat dit vagaal-nucleus solitarius-amygdala-insulair baansysteem verantwoordelijk is voor de viscerale feedback (Heilman, Bowers & Valenstein, 1993). |