![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
12 Cards in this Set
- Front
- Back
|
1.7 Functionele niveaus van het centraal zenuwstelsel
De opbouw van het centraal zenuwstelsel waarbij fylogenetisch jongere structuren een meer gespecialiseerde analyse brengen van inkomende informatie, gaf aanleiding tot een hiërarchisch model van het hersenfungeren.
Het meest eenvoudige model van deze benadering werd populair in het midden van de negentiende eeuw en stond centraal in de theorievorming van onder andere Spencer, MacLean en de globalist Hughlings-Jackson.
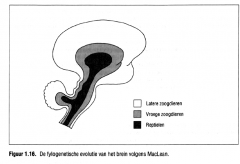
MacLean poneerde zijn 'triune brain ' waarbij het brein als
|
een drie-eenheid werd beschouwd met drie functionele niveaus die verwezen naar de gedragspatronen van reptielen, vroege zoogdieren en latere zoogdieren (Figuur 1.16.).
Met een toenemende fylogenetische ontwikkeling gaan zich eerst limbische structuren meer ontwikkelen waarna vooral een spectaculaire ontwikkeling van de corticale gebieden opvalt.
De onderliggende gedachte is onder meer dat de ontwikkeling van de nieu we (evolutief recente) hersengebieden een verfijnde controle mogelijk maakt op de fylogenetisch oudere en meer primitieve structuren (en de daaruit voortkomende driften). |
|
|
Met een toenemende fylogenetische ontwikkeling gaan zich eerst limbische structuren meer ontwikkelen waarna vooral een spectaculaire ontwikkeling van de corticale gebieden opvalt.
De onderliggende gedachte is onder meer dat |
de ontwikkeling van de nieu we (evolutief recente) hersengebieden een verfijnde controle mogelijk maakt op de fylogenetisch oudere en meer primitieve structuren (en de daaruit voortkomende driften). |
|
|
MacLean poneerde zijn 'triune brain ' waarbij het brein als een drie-eenheid werd beschouwd met drie functionele niveaus die verwezen naar de gedragspatronen van reptielen, vroege zoogdieren en latere zoogdieren (Figuur 1.16.). |
|
|
|
Klassiek werd de hiërarchische gelaagdheid van het zenuwstelsel onderzocht door bij proefdieren delen van het centraal zenuwstelsel te verwijderen en na te gaan welke functies dit gehavende zenuwstelsel nog kon uitvoeren.
Een dier waarbij hersenen en hersenstam werden verwijderd en dat enkel over het ruggenmerg beschikt, blijkt nog slechts tot zeer eenvoudige gedragingen in staat : |
de reflexen.
Hoewel het spinale dier niet kan staan noch zich spontaan kan bewegen, reageert het op reflexmatige wijze op zintuiglijke prikkels met onder meer strekbewegingen, terugtrekreflexen en krabben. |
|
|
Onderzoek bij dieren:
l h d g |
• het laag gedecerebreerde dier
|
|
|
Bij het laag gedecerebreerde dier werd naast de medulla spinalis ook het myelence phalon en metencepbalon intact gelaten.
Dit dier beschikt dus over verschillende myelencepbale en pontiene celgroepen die verantwoordelijk zijn voor vitale autonome functies, een deel van de formatio reticularis en de input van verschillende craniale zenuwen. Ook het cerebellum is aanwezig.
Gedragsanalyse toont dat deze dieren |
inactief zijn en hun lichaamstemperatuur slecht kunnen controleren. Ze vertonen wel slikbewe gingen en kunnen in de mond aangeboden voedsel tot zich nemen. Wanneer ze voldoende worden gestimuleerd, kunnen ze zich onhandig verplaatsen en vertonen ze rigide emo tioneel gedrag zoal grommen, bijten en blazen.
De dieren blijken opmerkelijk stijf. De stijfheid wordt veroorzaakt door een meer dan normale spanning van de spieren die normaal dienen om een opgerichte houding aan te nemen, en wordt gedecerebreerde rigidüeit genoemd. De dieren vertonen een aantal posturale (houdings)reflexen en een slaap-waakpatroon. |
|
|
In het hoog gedecerebreerde dier wordt ook het mesencephalon met tectum en tegmentum intact gelaten.
Het dier beschikt over de colliculi superior (visus) en inferior (auditus) en een aantal bijkomende motorische kernen. Dit dier vertoont evenmin spontane activiteit maar reageert op |
eenvoudige auditieve en visuele stimulatie en vertoont automatische (kauwen, pelsverzorging) en geïsoleerde willekeurige gedragingen zoals stappen, draaien , springen en klimmen wanneer het (voldoende) wordt gestimuleerd.
De componenten van de willekeurige bewegingen blijken dus aanwezig, maar worden niet in een context (bijvoorbeeld voor het vinden van voedsel) gebruikt. |
|
|
Bij het diencephale dier werd ook hypothalamus en thalamus intact gelaten.
Het dier is hyperactief en vertoont veel en spontane willekeurige bewegingen maar deze zijn niet adequaat. Het emotionele gedrag is complex, maar overdreven en weinig doelgericht ('sbam rage', zie hoofdstuk 'Emotioneel gedrag').
De hypothalamus zorgt dit keer voor
|
een adequate beheersing van de lichaamstemperatuur, maar het eet- en drinkgedrag blijft mede door de motorische hyperactiviteit ondermaats.
Het diencephalon biedt blijkbaar een activerende of motiverende bijdrage tot het gedrag, hoewel deze motivatie weinig doelgericht is. |
|
|
Bij het gedecorticeerde dier werd enkel de neocortex verwijderd.
Het dier beschikt over diencephalon en basale ganglia. Opvallend is dat in dit dier willekeurige en automatische bewegingen voldoende op elkaar afgestemd zijn zodat het dier in een eenvoudige omgeving weet te overleven (adequaat eet- en drinkgedrag).
De hyperactiviteit is sterk gereduceerd en het gedrag is doelgerichter. Toch vertoont het gedrag van deze dieren een aantal |
compulsieve kenmerken en blijken de gedragingen niet altijd even adequaat of goed uitgevoerd.
Ook bet emotioneel gedrag bleek niet altijd juist afgesteld op de stimuli die niet altijd correct worden geïnterpreteerd (zie Klüver-Bucysyndroom in het hoofdstuk 'Emotioneel gedrag').
De intacte neocortex in het normale dier blijkt vooral in te staan voor de fijne afstemming van doelgericht gedrag op een complexe omgeving. |
|
|
De resultaten van deze experimenten beklemtonen een |
hiërarchische structuur van het zenuwstelsel.
Hogere (fylogenetisch jongere) structuren zorgen voor een toenemende verfijning (controle) van het gedrag op de eisen van een complexe omgeving.
Ook de progressieve analyse van zintuiglijke informatie suggereert een hiërarchische organisatie waarbij een prikkel via het zintuiglijk baansysteem naar de thalamus wordt gevoerd en vanuit de thalamus naar de neocortex waar de prikkel na analyse in primaire, secundaire en tertiaire areas aanleiding geeft tot een respons die wordt opgebouwd in de motorische area.
De realiteit is echter veel ingewikkelder. |
|
|
De realiteit is echter veel ingewikkelder.
Verschillende thalamische kernen ontvangen zintuiglijke informatie en zenden die niet enkel naar de primaire cortex areas (hoewel dit wel de belangrijkste projectie is) maar ook naar verschillende secundaire areas.
Bovendien projecteren corticale areas terug naar de tbalamische kernen.
Dit model suggereert |
een parallelle verwerking van de informatie waarbij verschillende zones van de cortex verantwoordelijk zijn voor bepaalde aspecten van de stirnulusverwerking.
Deze hypothese wordt door klinische observaties bevestigd.
|
|
|
Specifieke corticale laesies laten soms heel specifieke gedragsstoornissen zien waarbij slechts bepaalde aspecten van een cognitieve functie worden getroffen zoals bij prosopagnosie, de onmogelijkheid om bekende aangezichten te herkennen.
Een dergelijke corticale organisatie impliceert tevens dat |
het moeilijker wordt een cognitieve functie, zoals bijvoorbeeld visuele waarneming, in de hersenen te lokaliseren vermits bepaalde aspecten van de visuele waarneming in de occipitale regio worden uitgevoerd en andere in de temporale of pariëtale regio. |