![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
57 Cards in this Set
- Front
- Back
|
acyl-CoAsynthetases |

catalyze the formation of athioester linkage between the fatty acid carboxyl group andthe thiol group of coenzyme A to yield a fatty acyl–CoA -34 kJ/mol |
|
|
Processing of dietary lipids in vertebrates (diagrams) |
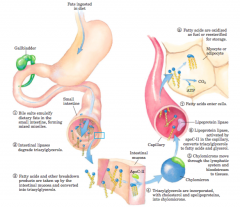
|
|
|
lipoprotein lipase |
activated by apoC-II, hydrolyzestriacylglycerols to fatty acids and glycerol |
|
|
perilipins |
a family of proteins that restrict access to lipiddroplets, preventing untimely lipid mobilization |
|
|
Cyclic AMP–dependent protein kinase (PKA) |
phosphorylates perilipin A phosphorylates hormone-sensitive lipase, doublingor tripling its activity, but the more than 50-fold increasein fat mobilization triggered by epinephrine is dueprimarily to perilipin phosphorylation |
|
|
Mobilization of triacylglycerols stored in adipose tissue (diagram and steps) |
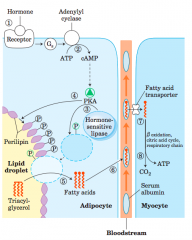
When low levels of glucose in the blood trigger the release of glucagon 1 the hormone binds its receptor in the adipocyte membrane 2 stimulates adenylyl cyclase, via a G protein, to produce cAMP. Thisactivates PKA, which phosphorylates 3 the hormone-sensitive lipaseand 4 perilipin molecules on the surface of the lipid droplet. Phosphorylationof perilipin permits hormone-sensitive lipase access to the surfaceof the lipid droplet, where 5 it hydrolyzes triacylglycerols to freefatty acids. 6 Fatty acids leave the adipocyte, bind serum albumin inthe blood, and are carried in the blood; they are released from the albuminand 7 enter a myocyte via a specific fatty acid transporter. 8 Inthe myocyte, fatty acids are oxidized to CO2, and the energy of oxidationis conserved in ATP, which fuels muscle contraction and other energy-requiringmetabolism in the myocyte. |
|
|
Entry of glycerol into the glycolytic pathway (diagram) |
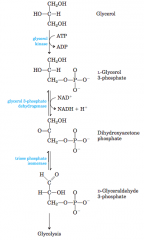
|
|
|
acyl-carnitine/carnitine transporter (diagram) |
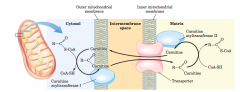
|
|
|
carnitine acyltransferaseII |
This isozyme, located on the inner face ofthe inner mitochondrial membrane, regenerates fattyacyl–CoA and releases it, along with free carnitine, intothe matrix Carnitine reenters the intermembranespace via the acyl-carnitine/carnitine transporter |
|
|
Coenzyme A (Co-A) |
in the mitochondrial matrix, largely used in oxidative degradation of pyruvate,fatty acids, and some amino acids in cytosol, used in the biosynthesis of fatty acids. Can be used for membrane lipid synthesis or can bemoved into the mitochondrial matrix for oxidation andATP production. |
|
|
Chylomicrons |
deliver triacylglycerols to tissues,where lipoprotein lipase releases free fatty acidsfor entry into cells. Triacylglycerols stored inadipose tissue are mobilized by a hormone-sensitivetriacylglycerol lipase. The released fatty acids bindto serum albumin and are carried in the blood tothe heart, skeletal muscle, and other tissues thatuse fatty acids for fuel. |
|
|
Molecular structure of a chylomicron (diagram) |
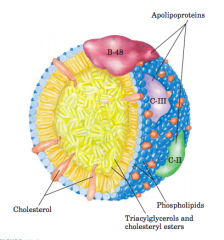
The surface is alayer of phospholipids, with head groups facing the aqueous phase.Triacylglycerols sequestered in the interior (yellow) make up more than80% of the mass. Several apolipoproteins that protrude from the surface(B-48, C-III, C-II) act as signals in the uptake and metabolism ofchylomicron contents. The diameter of chylomicrons ranges fromabout 100 to 500 nm. |
|
|
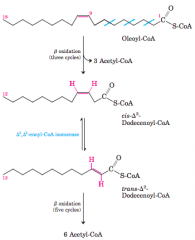
3 Stages of fatty acid oxidation (diagram) |
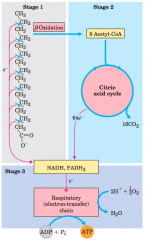
Stage 1: A long-chain fattyacid is oxidized to yield acetyl residues in the form of acetyl-CoA. Thisprocess is called beta oxidation. Stage 2: The acetyl groups are oxidizedto CO2 via the citric acid cycle. Stage 3: Electrons derived from theoxidations of stages 1 and 2 pass to O2 via the mitochondrial respiratorychain, providing the energy for ATP synthesis by oxidativephosphorylation. |
|
|
Conversion of a fatty acid to a fattyacyl–CoA (diagram) |
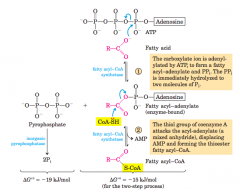
The conversion is catalyzed by fatty acyl–CoA synthetaseand inorganic pyrophosphatase. Fatty acid activation by formation ofthe fatty acyl–CoA derivative occurs in two steps. The overall reactionis highly exergonic |
|
|
Beta Oxidation Pathway Reactions (diagram) |
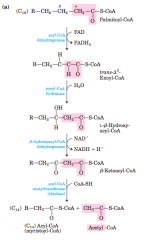
In each pass through thisfour-step sequence, one acetyl residue (shaded in pink) is removed inthe form of acetyl-CoA from the carboxyl end of the fatty acyl chain—in this example palmitate (C16), which enters as palmitoyl-CoA |
|
|
In one pass through the Beta-oxidation sequence.... |
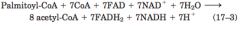
one moleculeof acetyl-CoA, two pairs of electrons, and four protons(H") are removed from the long-chain fattyacyl–CoA, shortening it by two carbon atoms |
|
|
enoyl-CoA isomerase |
repositions the doublebond, converting the cis isomer to a trans isomer, a normal intermediatein beta oxidation |
|
|
Oxidation of a polyunsaturated fatty acid (diagram) |
Oxidation requires asecond auxiliary enzyme in addition to enoyl-CoA isomerase: NADPH dependent2,4-dienoyl-CoA reductase |
|
|
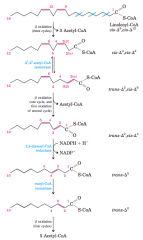
Oxidation of a monounsaturated fatty acid (diagram) |
|
|
|
Linking number (diagram and definition) |
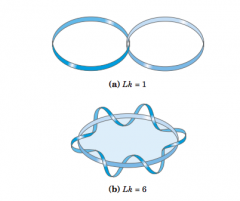
topologicalproperty of double-stranded DNA, because itdoes not vary when the DNA is bent or deformed, aslong as both DNA strands remain intact number of timesthe twisting strand penetrates this surface provides a rigorous definitionof linking number. |
|
|
topoisomerases |
enzymes that increase ordecrease the extent of DNA underwinding; change linking number |
|
|
Type Itopoisomerases |
act by transiently breaking one of thetwo DNA strands, passing the unbroken strand throughthe break and rejoining the broken ends they change Lkin increments of 1 |
|
|
Type II topoisomerases |
break both DNA strands and change Lk in increments of 2 |
|
|
Mechanism for topoisomerase Type 1 (diagram) |
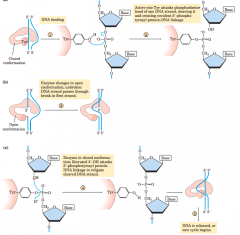
|
|
|
Mechanism for type 2 topoisomerases |
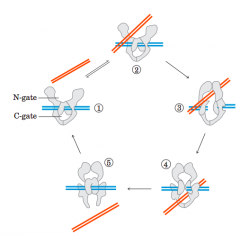
1 The multisubunitenzyme binds one DNA molecule (blue). Gated cavities above andbelow the bound DNA are called the N-gate and the C-gate. 2 Asecond segment of the same DNA molecule (red) is bound at theN-gate, and 3 is trapped. Both strands of the first DNA are nowcleaved and 4 thesecond DNA segment is passed through the break. 5 The brokenDNA is relegated, and the second DNA segment is released through theC-gate. Two ATPs are bound and hydrolyzed during this cycle |
|
|
histones |
package and order the DNA into structuralunits called nucleosomes |
|
|
Levels of organization of DNA (diagram) |
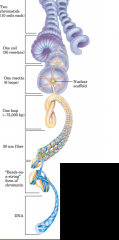
Nucleosome is DNA-wrapped histone |
|
|
DNA ligases |
After an RNA primer is removed and thegap is filled in with DNA, a nick remains in the DNA backbonein the form of a broken phosphodiester bond. Thesenicks are sealed by DNA ligases |
|
|
Dam methylase |
methylates DNA at the N6 position of all adenineswithin (5')GATC sequences |
|
|
Methylation and mismatch repair |
distinguish parent (template) strands from newlysynthesized strands in E. coli DNA, a function that is critical tomismatch repair The methylation occurs at the N6 ofadenines in (5')GATC sequences. For a short periodfollowing replication,the template strand ismethylated and thenew strand is not. After a few minutesthe new strand ismethylated and thetwo strands can nolonger be distinguished. |
|
|
DNA glycosylases |
recognize particularlycommon DNA lesions andremove the affected base by cleaving the N-glycosylbond. Uracil DNA glycosylases, for example, found inmost cells, specifically remove from DNA the uracilthat results from spontaneous deamination of cytosine.Mutant cells that lack this enzyme have a high rate ofG=C to AUT mutations. |
|
|
Base-excision repair pathway(diagram) |
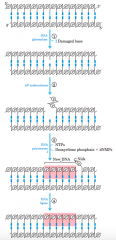
1 A DNA glycosylase recognizes a damaged base and cleaves betweenthe base and deoxyribose in the backbone. 2 An AP(abasic site) endonucleasecleaves the phosphodiester backbone near the AP site. 3 DNApolymerase I initiates repair synthesis from the free 3' hydroxyl at thenick, removing (with its 5! --> 3! exonuclease activity) and replacing aportion of the damaged strand. 4 The nick remaining after DNA polymeraseI has dissociated is sealed by DNA ligase. |
|
|
Repair of pyrimidine dimers with photolyase(diagram-mechanism) |
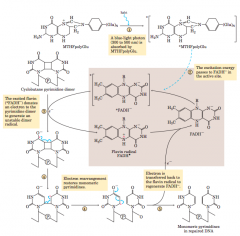
|
|
|
Catalytic mechanism of RNA synthesis by RNA polymerase (diagram) |
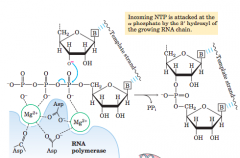
|
|
|
promoters |
specific sequences inthe DNA which direct the transcriptionof adjacent segments of DNA (genes). |
|
|
consensus sequence |
certain nucleotidesthat are particularly common at each position (-10 and -35) of the promoter region Mutations that affect the function of a givenpromoter often involve a base pair in these regions. Variationsin the consensus sequence also affect the efficiencyof RNA polymerase binding and transcription initiation |
|
|
Footprinting |
technique derived from principlesused in DNA sequencing, identifies the DNA sequencesbound by a particular protein |
|
|
the sigma cycle(diagram) |
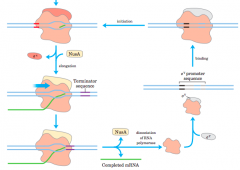
|
|
|
processivity |
enzyme's ability to catalyze "consecutive reactions without releasing its substrate" DNA polymerases associated with DNA replication tend to be highly processive, while those associated with DNA repair tend to have low processivity |
|
|
RNA polymerase I (Pol I) |
responsible for thesynthesis of only one type of RNA, a transcript called preribosomalRNA (or pre-rRNA) |
|
|
RNA polymeraseII (Pol II) |
synthesis of mRNAs and some specializedRNAs |
|
|
RNA polymerase III (Pol III) |
makes tRNAs, the 5SrRNA, and some other small specialized RNAs |
|
|
splicing |
introns are removed fromthe primary transcript and the exons are joined to forma continuous sequence that specifies a functionalpolypeptide |
|
|
5' cap |
eukaryotic mRNAs have a 5' cap, a residue of 7-methylguanosine linked to the 5'-terminal residue of themRNA through an unusual 5',5'-triphosphate linkage. The 5' cap helps protect mRNA from ribonucleases.It also binds to a specific cap-binding complexof proteins and participates in binding of the mRNAto the ribosome to initiate translation |
|
|
Splicing mechanism of groupI introns(diagram) |
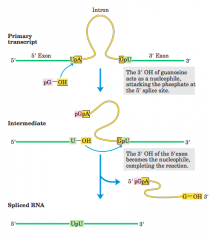
|
|
|
Transesterification reaction (diagram-mechanism) |
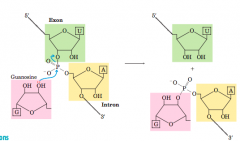
|
|
|
poly(A) tail |
At the 3' end, most eukaryotic mRNAs have a string of80 to 250 A residues Thistail serves as a binding site for one or more specific proteins. Protects mRNA from enzymatic destruction. Note: Many bacterial mRNAs also acquire poly(A) tails, butthese tails stimulate decay of mRNA rather than protectingit from degradation |
|
|
introns |
integral to gene expression regulation spliced out during RNA processing |
|
|
Creation of RNA cap (diagram-reactions) |
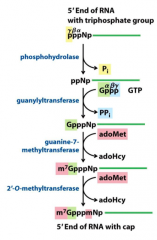
|
|
|
spliceosome |
the introns undergosplicing by the same lariat-forming mechanism asthe group II introns. The spliceosome is made up of specializedRNA-protein complexes, small nuclear ribonucleoproteins(snRNPs, often pronounced “snurps”). Each snRNP contains one of a class of eukaryotic RNAs,100 to 200 nucleotides long, known as small nuclearRNAs (snRNAs). Five snRNAs (U1, U2, U4, U5, andU6) involved in splicing reactions are generally found inabundance in eukaryotic nuclei. The RNAs and proteinsin snRNPs are highly conserved in eukaryotes fromyeasts to humans |
|
|
Splicing mechanism of group II introns(diagram) |
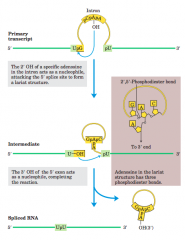
|
|
|
Termination codons |
UAA, UGA, UAG |
|
|
Initiation codon |
AUG |
|
|
wobble hypothesis |

|
|
|
anticodon |
Transfer RNAs base-pair with mRNA codons at athree-base sequence on the tRNA |
|
|
termination factors |
the proteins RF-1, RF-2, and RF-3—contribute to (1) hydrolysis of the terminal peptidyltRNAbond; (2) release of the free polypeptide and thelast tRNA, now uncharged, from the P site; and (3) dissociationof the 70S ribosome into its 30S and 50S subunits,ready to start a new cycle of polypeptide synthesis(Fig. 27–31). RF-1 recognizes the termination codonsUAG and UAA RF-2 recognizes UGA and UAA. EitherRF-1 or RF-2 (depending on which codon is present)binds at a termination codon and induces peptidyltransferase to transfer the growing polypeptide to a watermolecule rather than to another amino acid |
|
|
elongation factors |
elongation requires three soluble cytosolicproteins (EF-Tu, EF-Ts, andEF-G in bacteria), and (4) GTP |