Reading...
![]()
Play button
![]()
Play button
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
200 Cards in this Set
- Front
- Back
- 3rd side (hint)
|
reversal potential
|
also known as the nernst potential
the driving force of an ion through a channel essentially I=G(V-E) I is current-can be meausured using a voltage clamp V=voltage across membrane...meausred microelectrodes E=-60/z *log(ion in/ion out conc) |
|
|
|
PSC
|
postsynaptic current
|
|
|
|
PSP
|
postsynaptic potential
on postsynaptic cell graded potentials |
|
|
|
EPP
|
macroscopic current resulting from the summed opening of many ion channels
end plate is always summed value of many channels |
|
|
|
EPSP
|
excitatory post synaptic potential
|
|
|
|
IPSP
|
inhibitory post synaptic potential
|
|
|
|
What determines postsynaptic excitation and inhibition?
draw a picture |
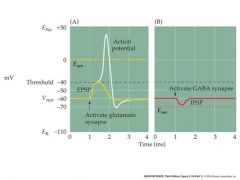
1)nernst potential
...if more positive than threshold, will be excitatory ...if less positive, will be inhibitory glutamat channels are non-selective cation GABA channels are chloride inhibitory |
|
|
|
what contains graded potentials?
what contains action potentials? |
graded: dendrites contain NT-gated channels
action: cell body + axon contains voltage-dependent channels |
|
|
|
draw an example of summation
|
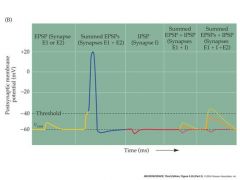
|
|
|
|
draw the summation pathway
|
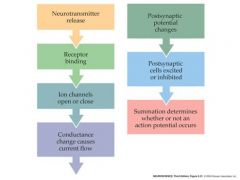
|
|
|
|
NT gated channels also called what?
talk about these |
ionotropic
NT-gated channels are also called “ionotropic” neurotransmitter receptors. Ionotropic receptors allow a NT to very rapidly change the membrane potential of the dendrites and cell body of postsynaptic cell, altering the frequency of a.p.’s In addition, if the NT-gated ionotropic receptor is permeable to calcium ions, these will diffuse into the postsynaptic cell and may stimulate enzymes and alter protein function or expression, acting as messenger molecules Ionotropic NT receptors are complex molecules – Made of several large subunits. The “nicotinic” Ach receptor at the neuromuscular junction has a relatively simple gating function – Only Ach required. |
|
|
|
draw ionotropic receptor
|
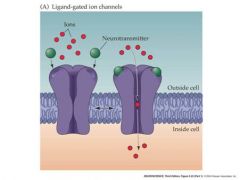
ligand gated receptors
|
|
|
|
draw acetycholine metabolism in cholinergic nerve terminals
what kind of receptor is this? |
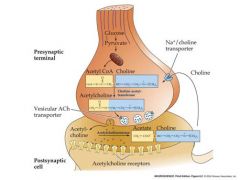
ionotropic
|
|
|
|
draw glutamate synthesis cycle
|
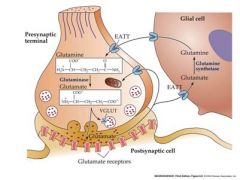
|
|
|
|
what are two examples of extremely simple receptors?
|
nicotinic ACh receptor
and the Glutamate AMPA receptor |
|
|
|
talk about the NMDA receptor
p |
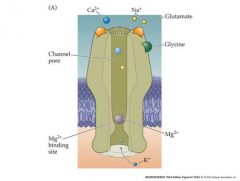
-depolarization forces Mg ion out of the channel and unblocks it....hyperpolarization blocks channel
if the cell is not previosly depolarized, there will be no response to glutamate |
N
Magnesium Depolarized And........glycine |
|
|
draw the EPSC projection of a NMDA channel
|
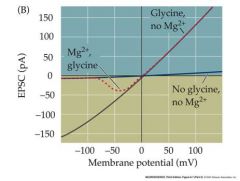
So, even if a synapse that releases glutamate is active, NMDA receptors will only be fully open if the cell is previously depolarized and the Mg block is therefore removed.
The NMDA receptor therefore creates a glutaminergic synapse that is only effective if the postsynaptic cell has been recently activated AND IS STILL DEPOLARIZED by another synapse – a mechanism useful for creating an association between two synaptic inputs to a neuron Involved in learning and memory at the cellular level |
|
|
|
what is interesting about a single NT such as glutamate?
|
can act on a diverse population of receptors to create postsynaptic potential with different characteristics and functions
|
|
|
|
2 inhibitory neurotransmitters
|
GABA and Glycine
|
|
|
|
synthesis and release of GABA
|
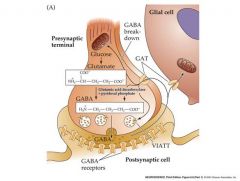
VIATT: vesicular inhibitory amino acid transporter
|
|
|
|
draw a membrane potential scheme of GABA inhibition
|
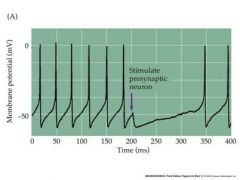
stimulation of presynaptic GABAergic interneuron
|
|
|
|
third of the synapses in the braine use what inhibitory neurotransmitter?
|
GABA
|
|
|
|
GABAa and GABAb...which one is ionotropic and which one is metabotropic?
|
ionotropic: GABAa
metabotropic: GABAb |
|
|
|
Draw the GABA receptor
|
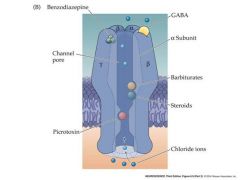
only gaba is needed to activate...all of the other subunits change the receptor in some manner
there are actually 2 sites for GABA to bind |
Gabby did drugs
|
|
|
are ionotropic NT receptors complex simple or complex proteins?
|
complex....multiple spanning subunit and usually more than one binding site for the NT
|
|
|
|
draw the nACh membrane spanning receptor
|
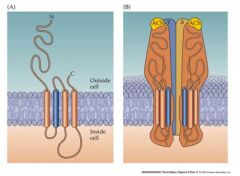
|
|
|
|
most important receptor in brain function
|
glutamate
|
|
|
|
describe architecture of ligand-gated receptors
p |
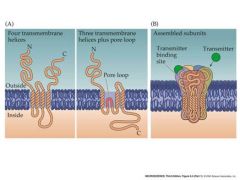
|
|
|
|
basics of metabotropic NT receptors
|
Metabotropic NT receptors
Gpro couples Initiate production of second messengers and acrivation of remote ion channels Act via GTP-binding proteins Potential for enormous signal amplification Slow response time |
|
|
|
very basic of G pro coupled receptor and posible response
|
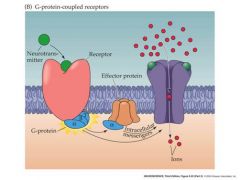
|
|
|
|
types of gtp-binding protein
p 2 types |
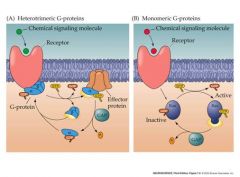
|
|
|
|
draw amplification scheme
|
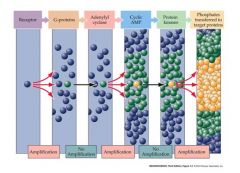
|
|
|
|
besides altering membrane potentials in postsynaptic cells, what else can NT's do?
|
In addition to altering the membrane potential of postsynaptic cells, by opening ion channels, NT’s can act to regulate the long-term activity of the postsynaptic cell.
This regulation can occur through the regulation of gene expression by a variety of possible pathways…. |
|
|
|
All of the classic second messenger pathways are available to NT’s via:
|
activation of metabotropic receptors
Activation of calcium-permeable ionotropic receptors leading to increased postsynaptic cellular calcium ion concentration. |
|
|
|
Talk about CREB
p |
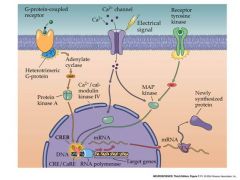
Phosphorylation of CREB allows it to bind co-activators:not shown.
This stimulates RNA polymerase to begin synthesis of RNA |
|
|
|
diagram and talk about long-term synaptic depression
|
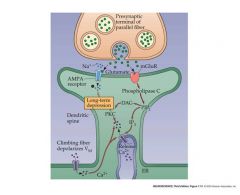
Activation of protein kinases by second messengers can also lead to phosphorylation of target proteins in the postsynaptic cell and medium-term changes in postsynaptic cell function.
These changes in function are often targeted towards proteins that regulate the sensitivity of the synapse to further stimulation by NT’s Thus stimulation of a synapse leads via this feedback to long term changes in the sensitivity of that synapse – a form of “learning”. For instance prolonged activity at synapses made with cerebellar neurons leads to a long-term reduction in the sensitivity of the post-synaptic receptors to NT’s – “long-term synaptic depression”. |
|
|
|
what does a cerebellar purkinje cell look like?
|
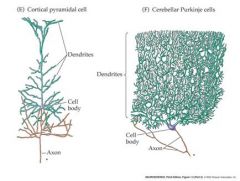
|
|
|
|
what can alter long-term strenght of pre-synaptic terminal?
|
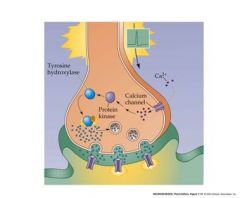
Calcium influx into the presynaptic terminal during prolonged stimulation can also result in protein phosphorylation and long-term changes in synaptic strength.
E.g. stimulation of tyrosine hydroxylase – an enzyme needed for biogenic amine synthesis. In this case, prior activity at the synapse results in an increase in long term synaptic strength – more amine NT is produced. |
|
|
|
plasticity basics
|
So chemical synapses allow both direct, immediate transmission of information via changes in membrane potential…..
Regulation of postsynaptic action potential frequency and more subtle, long-term regulation…. through intracellular messenger pathways, the synapses are rendered “plastic” – the strength of the connection depends on the amount of prior activity at a synapse. Plasticity is the building-block of learning and memory. |
|
|
|
draw a scheme of peptide neurotransmitter processing
|
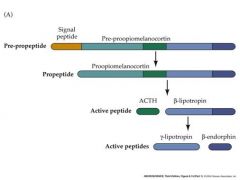
Pre-propeptide occurs in RER
Propeptide occurs in Golgi |
|
|
|
3 peptide categories with examples and types of amino acid properties
|
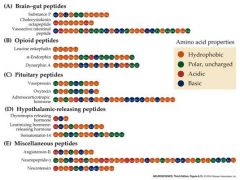
|
|
|
|
talk about opioid family of peptide NT's
|
Leu-Enkephalin, the dynorphins and the endorphins are members of the opioid family of peptide NT’s
They bind to the same postsynaptic receptor proteins that bind opium (morphine) and synthetic opiates (fentanyl) Opioid peptides are widely distributed in the presynaptic terminals of neurons throughout the brain – often co-localized with small-molecule NT’s like GABA or 5H-T When injected into the brain opioid peptide NT’s act as analgesics and depressants Opioid peptide generally act through G-protein linked receptors, not ionotropic receptors -amplified long term effects on postsynaptic cell metabolism and function |
|
|
|
2 unconventional NT's
|
Endocannabinoids
Gases – NO (nitric oxide) |
|
|
|
marijuana plant name
|
Cannabis sativa
|
|
|
|
active component of marijuana
|
THC
tetrahydrocannabinol |
|
|
|
areas of the brain that marijuana affects
|
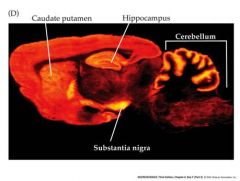
1)Substantia nigra
2)Caudate putamen 3)Hippocampus 4)Cerebellum |
|
|
|
Endo-cannabinoids
|
Endo-cannabinoids
- bind to cannabinoid receptor proteins – the proteins that bind THC, the active ingredient of marijuana They are fatty acids with polar head groups Produced in the cells by enzymes that are activated by Ca2+. Diffuse through membrane onto target cell Participate in synaptic regulation, rather than actual transmission. E.g. retrograde inhibition of inhibitory synapses in the cerebellum and hippocampus -one of their main functions is to regulate the release of GABA at inhibitory terminals |
|
|
|
retrograde signalling
|
In neuroscience, retrograde signaling is a phenomenon in which a signal travels from a postsynaptic neuron to a presynaptic one. Endocannabinoids are known to play a role in retrograde signaling, with the molecules being synthesized postsynaptically and acting presynaptically. Retrograde signaling may also play a role in long-term potentiation, a proposed mechanism of learning and memory, although this is controversial
|
|
|
|
GABA function
|
In vertebrates, GABA acts at inhibitory synapses in the brain by binding to specific transmembrane receptors in the plasma membrane of both pre- and postsynaptic neuronal processes. This binding causes the opening of ion channels to allow the flow of either negatively charged chloride ions into the cell or positively charged potassium ions out of the cell. This action results in a negative change in the transmembrane potential, usually causing hyperpolarization
|
|
|
|
talk about endocannibinoid-mediated retrograde control of GABA release
|
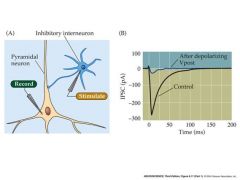
-depolarization of the postsynaptic neuron causes increases in calcium levels
-increased calcium levels cause synthesis and release of endocannabinoids -retrograded from post synaptic cell to pre synaptic cell -Bind to CB1 receptors and inhibit the amount of GABA released when pre-synaptic cells have AP's |
|
|
|
talk about NO and its effects
|
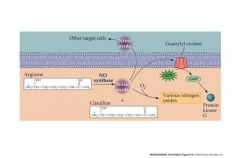
Produced by a Ca2+ - activated enzyme, NO-synthase, present in some neurons and other cells
Diffuses across to other neurons to stimulate enzymes produce the messenger cGMP in the “postsynaptic” cell Well-known role in inducing muscle relaxation |
|
|
|
somatosensation
|
Somatosensation is a sensory modality that covers
Touch and vibration Pain , temperature (considered by some to be separate) Proprioception - information from joint and muscle receptors – used to control muscle movement, not perceived Mechanosensory information from receptor cells in the viscera |
|
|
|
ganglion
|
collection of neuronal cell bodies outside of the CNS
|
|
|
|
medulla oblongata
|
The medulla oblongata is the lower portion of the brainstem. It deals with autonomic functions, such as breathing and blood pressure. The cardiac center is the part of the medulla oblongata responsible for controlling the heart rate.
|
|
|
|
dorsal root ganglion
|
little notches of ganglions off of spinal cord
|
|
|
|
preganglion fibers
|
originate in the brain or spinal cord
|
|
|
|
describe the path of mechanosensation and also to a lesser degree of pain
|
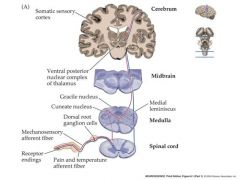
Machanosensation begins with a mechanosensory receptor neuron.
This neuron typically has: Its cell body in the dorsal root ganglion of the spinal cord. Its dendrites in layers of the skin (or viscera) An axon that travels up the spinal cord to make excitatatory synaptic contact with “second order” neurons in the medulla The second order mechanosensory cell, with dendrites and cell body in the cuneate or gracile nucleus of the medulla, sends an axon that terminates in the ventral posterior nuclear complex of the thalamus, making an excitatory synaptic connection with a “third order” neuron. The third order mechanosensory cell, with dendrites and cell body in the VPN thalamus, sends an axon that terminates in the post-central gyrus of the cerebral cortex, making contact with cerebral neurons. Thus the post central gyrus is the somosensory area of the cerebral cortex. Pain and temperature info. travels by a slightly different pathway to the somatosensory cortex ipsalateral side is the side that the nerve enters on....contralateral side is the oposite side |
|
|
|
dorsal
|
backside of spine, top part of brain
|
|
|
|
ventral
|
front side of spine, lower part of brain
|
|
|
|
Rostral
|
toward front of brain or toward the head
|
|
|
|
caudal
|
toward the feet or toward the rear of the brain
|
|
|
|
superior
|
straight up
|
|
|
|
inferior
|
straight down
|
|
|
|
posterior
|
backwards
|
|
|
|
anterior
|
forward
|
|
|
|
describe mechanotransduction
10/16 pictures if needed long explanation required |
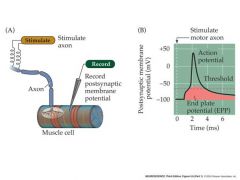
The transmission of sensory information begins with the transduction of a mechanical stimulus at the skin into an increase in the frequency of action potentials traveling through the mechanosensory neuron
The plasma membrane of the dendrites contain mechanosensitive channels, mostly permeable to sodium ions. Mechanotransduction begins when the dendrites of the mechanosensory neuron, under the skin, are distorted by a mechanical stimulus. This stresses the cell membrane and opens mechanosensitive channels – which are cation non-selective. Sodium ions flow through the channels and depolarize the dendrites of the mechanosensitive neuron The resulting depolarization is called the “generator” or “receptor” potential. It is a graded {passive} potential which increases in amplitude with stimulus strength as more channels open). It is similar in its nature to the EPP or PSP If the stimulus is strong enough, the generator potential will grow to exceed threshold and an action potential will be produced. The action potential will propagate down the axon to the spinal cord and beyond. If the stimulus is maintained, a train of action potentials will be generated in the mechanosensory neuron. The frequency of action potentials in the train will be proportional to the stimulus strength – up to a maximum frequency, when the neuron “saturates”. Thus information about stimulus strength is encoded in terms of the frequency of action potentials, Between threshold and saturation lies a dynamic range – a range of stimulus strength within which the a.p. potential activity within a neuron reflects stimulus strength. The action potential travels down the axon, the generator potential, Being passive, does not spread far down the axon. So only the Action potential arrives at the spinal cord |
|
|
|
draw action potential frequency saturation curve
|
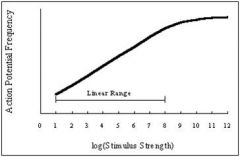
all neurons have spontaneeous background
spontaneous background is the flat part of the curve Spontaneous background is random |
|
|
|
what is an important feature that varies from one mechanosensory neuron to another?
|
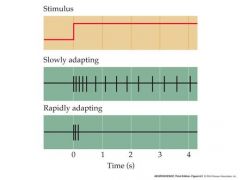
An important feature that varies from one mechanosensory neuron to another is the degree to which the neuron’s response tracks sustained stimulation.
For some mechanosensory neurons, the a.p. frequency drops off during prolonged stimulation – the neuron “adapts”. This means that the neuron responds best to phasic, or rapidly changing stimuli and not to steady stimuli. Other neurons respond steadily to prolonged stimulation – their response is said to be “tonic” Adaptation represents an important early step in processing sensory information – it tunes some receptors to rapidly changing, high frequency stimuli like vibration and it reduces the impact of unchanging signals that are of less “interest”. Something that wants to detect vibrations is said to be phasic |
|
|
|
talk about skin mechanoreceptors
|
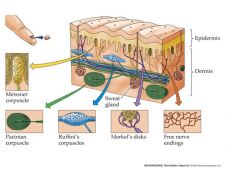
Meissner corpuscle: Touch Phasic
Pacinian Corpuscle: Vibration Phasic Ruffini's Corpuscles: Stretching Tonic Merkel's disks: Touch Pressure tonic Free Nerve endings: pain, temp, crude touch tonic, hig threshold |
|
|
|
what are 2 proprioceptrive sensory structures and describe them
|
muscle spindle and golgi tendon organ
muscle spindle: detects muscle length and initiates the stretch reflex golgi tendon organ: receptor that detects tension in tendon Axons from proprioceptive sensory structures in muscles, joints and tendons send branches via the dorsal root ganglion to the spinal cord and to the medulla (and thence to the thalamus and the somatosensory cortex) |
|
|
|
how do mechanoreceptors from upper and lower body differ?
|
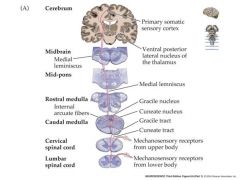
mechanosensory nerves from face also differ and they will be shown in another slide
|
|
|
|
mechanosensory receptor from face path
|
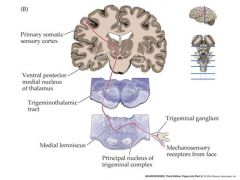
differ from mechanoreceptors from upper and lower body
enters at the midbrain |
|
|
|
what do proprioceptors do?
|
provide information about the position of the limbs and other body parts in space
|
|
|
|
diagram showing the somatic sensory numbers, and big picture of secondary somatic sensory cortex
|
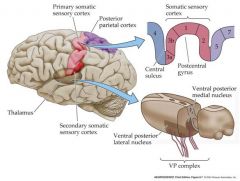
|
|
|
|
what are dermatomes....what's special about them?
|
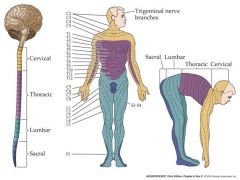
A closer inspection of this crude division into pathways conveying mechanosensory information from the upper, lower bodies and the face reveals more detail
In fact, sensory neurons within each dermatome of the skin surface “projects to” to a particular area within the somatosensory cortex (S1). Sensory neurons with dendritic fields in neighboring areas of the skin surface project to neighboring areas of the cortical surface. Creates a “somatotopic map” of the body surface which devotes more cortical surface to areas of the skin with greater receptor density. |
|
|
|
what makes up the brainstem?
|
midbrain, pons, and the medulla
|
|
|
|
draw a somatotopic map of humans
|
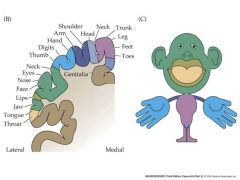
|
|
|
|
corical columns
|
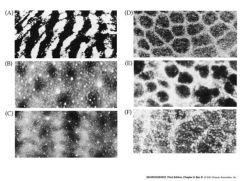
Within each little section of the cerebral cortex – dealing with a particular part of the skin - are columns of neurons with overlapping receptive fields that are arranged perpendicular to the cortical surface
each column of neurons tends to respond to a similar kind of sensory stimulus (e,g, vibration, touch, pressure etc.) Sometimes these columns are grouped to create a little columnar module that might, for instance, process information from an individual whisker on a rodent’s snout. These modules sometimes stain differently from surrounding tissues and so can be visualized. |
|
|
|
basics of pain perception and experiment that demonstrated that nociception involved specialzed neurons
|
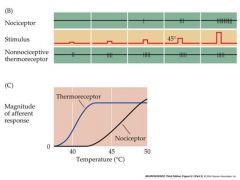
Is a sensory modality that is related to, but separately perceived from mechanoreception.
The perception of pain does not simply arise from saturation of the response of mechanoreceptor or thermoreceptor neurons by intense stimuli. Rather…… Painful stimuli are detected by specialized high-threshold sensory neurons called nociceptors. The nociceptive pathway is a sub-division of the somatosensory pathway, involving similar, but distinct cell types stimulus is a heat stimulus on the fingers |
|
|
|
pain perception also called?
|
nociception
|
|
|
|
phases of pain?
|
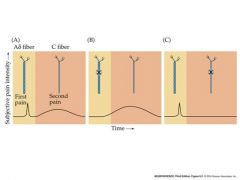
1)rapid sharp pain to spinal cord
2)later dull burning pain B and C represent blocking one of the axons so that the pain is not transmitted. |
|
|
|
active ingredient of chilis
picture of receptor |
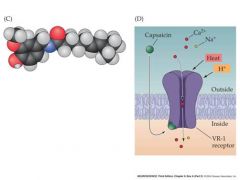
A subset of the small, unmyelinated “C” fibers
have non-specific cation channels in their dendrites that can be opened by Capsaicin – the active ingredient of chile peppers |
|
|
|
contralateral
|
opposite side
|
|
|
|
ipsilateral
|
same side
|
|
|
|
hemisection
|
damage to one half of the spinal cord...remember spinal cord has 2 sides of dorsal root ganglions
|
|
|
|
what would happen if a hemisection of the T10 spinal cord occured?
|
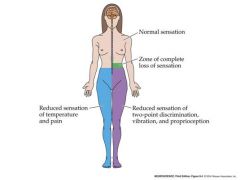
due to the fact that the nerves from pain and mechanoreception change sides
|
|
|
|
pain and temperature pathway in the body
|
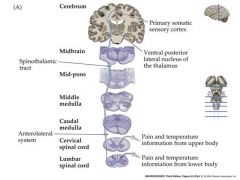
pathway crosses over immediately and travels up contralateral spinal cord
|
|
|
|
referred pain...2 examples
2 weird examples also |
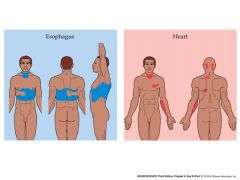
Because information from visceral pain sensory neurons is merged with those from
the skin in the CNS, pain in the viscera is often “referred” to the skin “Referred pain” brings up an important point concerning the functioning of somatotopic maps. Namely…… LABELED LINES OF INFORMATION FLOW IN THE CNS Neurons in CNS pathways are “labeled” in terms of the perceptions they evoke. It doesn’t matter how they are stimulated originally – neurons in a particular pathway will evoke the perception that they are “labeled” with. In the case of “referred pain”, when neurons in the CNS that are labeled as receiving information from the skin are instead activated via visceral sensory neurons, what is perceived is still the “label” – pain on the skin surface. As another example, consider placing pressure on your eyeball. What you perceive is light – due to the modulation of the activity of retinal neurons by the pressure. The information from these neurons are “labeled” in the CNS as indicating changes in the intensity of light. Labeled lines are also responsible for the referred pain known as phantom limb syndrome – thought to be due to the continued spontaneous activity of central neurons that would normally have received information from that limb |
|
|
|
analgesics
|
compound that reduces pains intensity
|
|
|
|
peripheral sensitization of nociceptors
|
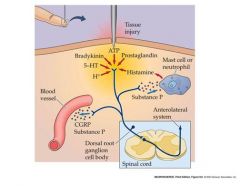
-creat a more painful feeling to help with healing and prevent further injury
-prostaglandins contribute to peripheral sensitization by binding to Gpro receptors that increase levels of cAMP...also reduce the threshold depolarization required for AP's other factors are responsible for vasodilation and swelling. |
|
|
|
pain perception overview and different pathways
|
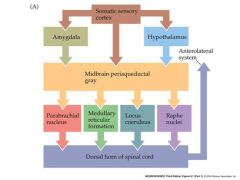
There is plenty of evidence that our perception of pain can be enhanced or depressed by the CNS
Ignoring pain during sports or battle Ability of mechanoreceptive stimuli to reduce perception of pain (acupuncture?) A variety of neuronal pathways have been defined in the CNS that modulate the perception of pain. These converge on the second order “projection” nociceptive neurons in the dorsal horn that transmit pain information up the anterolateral system to the medulla |
|
|
|
gate theory of pain
|
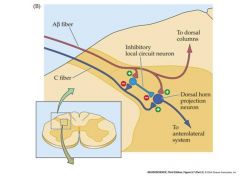
activation of mechanoreceptors can inhibit the transmission of ascending pain signals
if you hit your shin hard, it will feel better if you rub the overlying mechanoreceptors C fiber is the nociceptor A fiber is the mechanorecptor |
|
|
|
draw a scheme of nociceptor inhibition in the dorsal horn
|
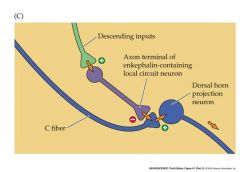
descending input is from Raphe nuclei
|
|
|
|
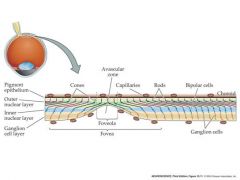
where is light focused in the eye?
anatomy of eye |
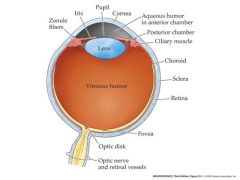
back, interior surface of the eyeball....which is covered with the retina-the neural layers of the eye
The retina has many layers of cells. Oddly, the photoreceptor cells (rods and cones) are at the back of the retina – facing towards the back of the eye - with their outer segments buried in a pigmented epithelium that absorbs scattered light. This is not a problem, since the layers of the retina lying over the photoreceptors are largely transparent and the visual tends to ignore any stationary shadows cast by blood vessels and nerves overlying the photoreceptors Iris: colored portion of eye pupil: black part of eye sclera: outermost tissue layer...the white stuff cornea: transparent tissue that permits light rays to enter the eye aqueous humor: fluid in the eye Optic disk: blind spot where nerves come from Fovea: center part of macula where acuity is the greatest |
|
|
|
types of eye defects
|
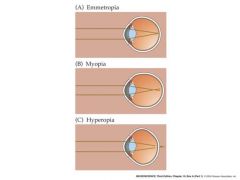
emmetropia: normal vision
myopia (near sighted): too much curvature of the cornea or by the eyeball being too long. image of distant object focuses in front of, rather than on, the retina hyperopia: farsighted, eyeball being too short or the refracting system too weak |
|
|
|
do photoreceptor cells have action potentials?
|
no
|
|
|
|
how does information flow from the retina?
|
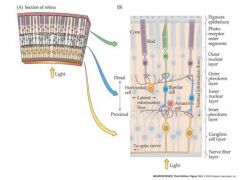
Light is absorbed by the photoreceptor (rod or cone) outer segments. This triggers a change in their membrane potential that is graded with light intensity (no a.p.s!)
The change in photoreceptor membrane potential is transmitted to the bipolar cell by the photoreceptor-bipolar cell synapse. As a result there is a change in bipolar cell membrane potential that is also graded with light intensity (no a.p.s!). The change in bipolar cell membrane potential is transmitted to the ganglion cell by the bipolar - ganglion cell synapse. Unlike the bipolar and photoreceptor cells, the ganglion cells has V-dependent sodium and potassium channels and so can generate action potentials As a result of illumination there is a change in ganglion cell membrane potential that results in a change in ganglion cell action potential frequency. The frequency of action potentials produced by the ganglion cell is graded with light intensity. The action potentials travel along the ganglion cell axons in the optic nerve to the brain |
|
|
|
unaccommodated and accommodated eye picture
|
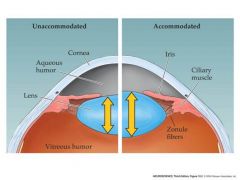
unaccommodated:cilliary muscles relaxed
accommodated: near objects...ciliary muscles contracted..reduces the tension in the zonule fibers allowing the lens to increase its curvature which allows for short distance.. When the cilliary is relaxed you can see long distance Circular muscles cause less eye dilation pupil muscles create more eye dilation |
|
|
|
structural difference between rods and cones
|
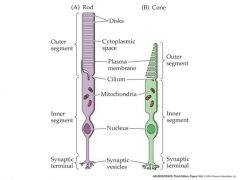
Rods are much more sensitive than cones. They generate large responses to single quanta of light but their response saturates in daylight.
Rods are slower than cones (linked to higher amplification). They take longer to respond and their response lasts longer – less temporal sensitivity. Rods are mostly found in the peripheral retina – the fovea contains only cones, packed at v. high density. Consequently, cones are good for capturing rapidly changing detailed images under daylight conditions. Rods are better for imaging in dim light, when amplification and quantum capture is important. Rods express only one rhodopsin (gene) which absorbs in the green part of the spectrum. A cone may express one of three different rhodopsin genes, absorbing in the red, green or blue part of the spectrum. So comparison of output from the three cone types can be used for color vision in daylight – no color vision from rods at night. |
|
|
|
intracellular recording from a single cone or rod stimulated with different amounts of light
|
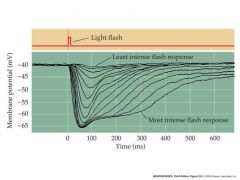
response is hyperpolarization
cell in dark is depolarized to -40mV In the light, it is hyperpolarized by decreasing permeability to calcium channels |
|
|
|
mechanism of visual transduction....picture, but basic
|
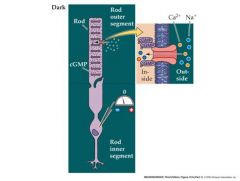
light causes the channel to close so that no sodium and calcium will enter.
|
|
|
|
picture of rod photoreceptor
|
Absorption of light causes the retinal to go from a cis conformation to a trans conformation
|
|
|
|
photoreceptor phototransduction pathway in rods
|
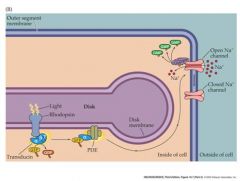
-change in retinal structure activates transducin, which in turn activates a PDC.
This PDE hydrolyzes cGMP, reducing its concentration in the outer segment and leading to the closure of channels in the outer segment membrane |
|
|
|
difference between rods and cones
|
Rods are much more sensitive than cones. They generate large responses to single quanta of light but their response saturates in daylight.
Rods are slower than cones (linked to higher amplification). They take longer to respond and their response lasts longer – less temporal sensitivity. Rods are mostly found in the peripheral retina – the fovea contains only cones, packed at v. high density. Consequently, cones are good for capturing rapidly changing detailed images under daylight conditions. Rods are better for imaging in dim light, when amplification and quantum capture is important. |
|
|
|
cone/rod spectrum in light
|
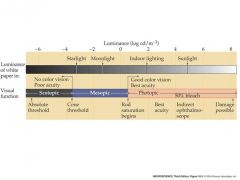
Scotopic: only rods
Once at Photopic, only cones make a difference because rods do not respond to intensity changes and thus are maxed out. mn: scott was a mess in his photos |
|
|
|
distribution of rods and cones in the human retina
|
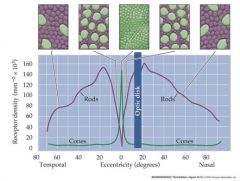
cones are present at a low density throughout the retina, with a sharp peak in the center of the fovea...rods a re present at high density throughout most of the retina, with a sharp decline in the fovea. rods are absent in the foveola
|
|
|
|
picture of fovea and foveola cells
|
|
|
|
|
how many rhodopsin genes do rods express and what do they absorb?
|
1 gene and green part of the spectrum
|
|
|
|
what spectrum can cones absorb in?
|
red
green blue |
|
|
|
is there any color vision output from rods at night?
|
no
|
|
|
|
color vision of cones and rod chart
|
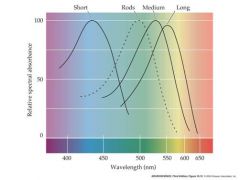
|
|
|
|
what is special about the red an green cone pigment?
|
genes lie close to each other on the X chromosome
Men with only one copy of these genes are more susceptible to colorblindness due to defects in these genes than are women (XX) who have two copies |
|
|
|
genetic alteration that causes deficiencies in color vision
|
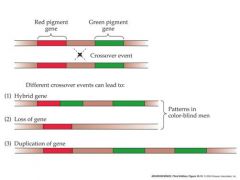
if there is alterations in the green gene, two of them can cause it to appear red
|
|
|
|
what happens to photoreceptors in response to light?
|
they hyperpolarize and decrease their neurotransmitter release
|
|
|
|
essentially what are ganglion cells good at?
|
they are sensitive to differences between the level of illumination that falls on the receptive field center and the level of illumination that falls on the surround....luminance contrast
|
|
|
|
receptive fields
|
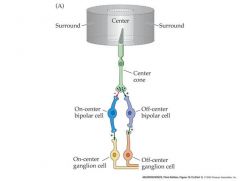
Responses of retinal ganglion cells (r.g.c’s) to light
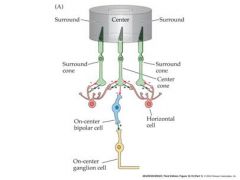
All r.g.c’s display some spontaneous a.p. activity – even in darkness. Many r.g.c’s adapt to prolonged stimuli (The change in a.p frequency decreases with time after light onset) Each retinal ganglion cell has a “receptive field” The area of the retina (or the equivalent section of visual space) within which light elicits a response from the cell. r.g.c receptive fields are always circular, and cover about 1 degree of visual space. Retinal ganglion cells fall into two types with slightly different receptive field properties: On-center r.g..c’s Respond to illumination of the center of the receptive field with an increase in a.p. frequency Off-center r.g..c’s Respond to illumination of the center of the receptive field with an decrease in a.p. frequency Illumination of the “surround” the area surrounding the center of the receptive field has the opposite effect to that of illuminating the center On-center r.g.c’s Respond to illumination of the surround of the receptive field with an decrease in a.p. frequency Off-center r.g.c’s Respond to illumination of the surround of the receptive field with an increase in a.p. frequency So r.g.c’s have “circular center-surround” receptive fields. Why? – what benefit is this to vision? It means that r.g.c’s are maximally stimulated by edges and lines, where intensity changes rapidly, and NOT by uniform illumination – they are “edge detectors” “Edge detection” is a form of spatial image filtering – allows more “bandwidth” to be allocated to edges and lines during transmission of information along the optic nerve. - and + indicate a sign conserving synapse or a sign-inverting synapse Remember cones and rods are hyperpolarized in response to light and release less glutamate...bipolar cells also release glutamate...glutamate is an excitatory NT On-center cells have a cGMP gated Na channel when bound to Glutamate...Off-center have a ionotropic receptor AMPA that causes the cell to depolarize to glutamate. |
|
|
|
draw the scheme for stimulating the center and surrounding of on an off center ganglion cells
|
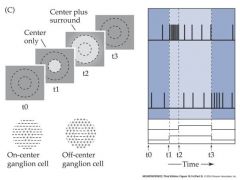
illumination of the surround antagonizes illumination of the center
|
|
|
|
draw a schem for the rate of discharge of an on-center ganglion to a "spot" of light....should include a curve
|
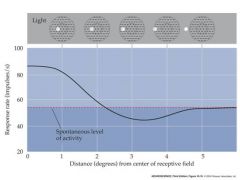
|
|
|
|
draw a scheme for the enhanced edge-detection by retinal ganglion cells
|
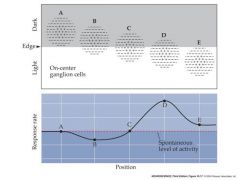
cells totally in light or darkhave little or no difference in response to uniform light or dark
|
|
|
|
what would AP's look like for a light spot in the center of an on and off center ganglion
|
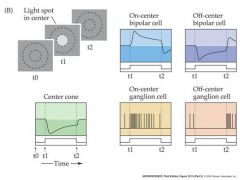
|
|
|
|
what would the voltage look like for a dark spot in the center of a ganglion cell
|
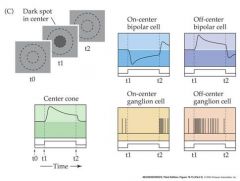
|
|
|
|
describe the circuitry responsible for generating the receptive field surround of an on-center retinal ganglion cell
|
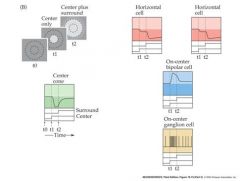
also look at picture on page 6 of 10/30 notes for a visual picture of how horizontal cellswork
Horizantal cell receives input via GAP junctions from photoreceptor terminals...When horizantal cells are depolarized they release the inhibitory nt GABA back on to retinal cells. When GABA is not released, the cone and rods become depolarized and reduce the light evoked response. Ultimately this reduces the firing rate of the on-center ganglion cells. |
|
|
|
What creates the center of the receptive field?
|
the vertical information frlowing from the center cone to bi polar cells>ganglion cells
off center and on center |
|
|
|
what creates the surrounding of the visual field?
|
lateral connections via horizontal cells
|
|
|
|
fovea
|
The fovea, also known as the fovea centralis, is a part of the eye, located in the center of the macula region of the retina. [1] [2] The fovea is responsible for sharp central vision (also called foveal vision), which is necessary in humans for reading, watching television or movies, driving, and any activity where visual detail is of primary importance.
|
|
|
|
Iris
|
Color part of the eye
|
|
|
|
Pupil
|
open black part of the eye
|
|
|
|
Cornea
|
clear transparent outer layer covering pupil and Iris
|
|
|
|
Sclera
|
White part of eye around cornea
|
|
|
|
fluid in the eye
|
vitreous humor
|
|
|
|
Retina
|
what receives light and detects an image
|
|
|
|
explain how horizontal cells work
|
horizontal cells receive input from photoreceptor terminals linked via GAP junctions
they influence the release of neurotransmitter directly onto photoreceptor terminals, regulating the amount of transmitter that the photoreceptors release onto bipolar cell dendrites Glutamate release from photoreceptor cells depolarizes horizontal cells....This causes GABA to be released by horizontal cells which has an inhibitory effect on photoreceptor cells. As a result, horizontal cells work to prevent changes in photoreceptor cells. Horizontal cells are the reason that there is a surround field around the center visual field. |
|
|
|
Draw the typical visual field and how it would be produces when it goes through the eye
|
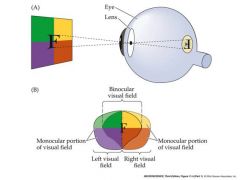
everything is inverted
top goes to bottom...right goes to left.... Likewise nasal visual field is actually temporal on the retina surface |
|
|
|
draw a left and right monocular visual field and how it would be produced on the retina
|
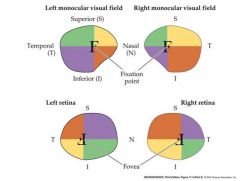
a
|
|
|
|
draw the visual field and how various points would be projected onto the retina....How would these lead to the brain?
|
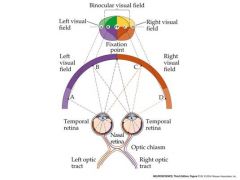
nasal-contralateral....crosses
temporal....ipsalateral....does not cross |
|
|
|
Draw the path of a signal after light is detected....trace the brain pathway
|
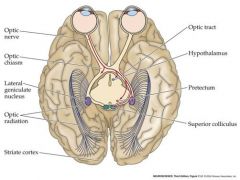
hypothalamus is involved in circadian rhythm: biological clock
Pretectum: reflex control of pupil and lens Superior colliculus: orienting the movements of head and eyes Major target is the Lateral geniculate nucleus MN: hippo protected the cool gent in the cort |
|
|
|
diencephalon
|
contains the thalamus and hypothalamus
|
|
|
|
the following part of the visual pathway is damaged, what would be the consequence?
right optic nerve optic chiasm right optic tract top part of right optic radiation all of right optic radiation |
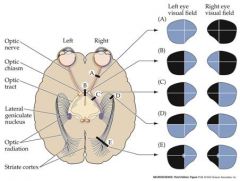
|
|
|
|
How is the visual field represented in the brain
|
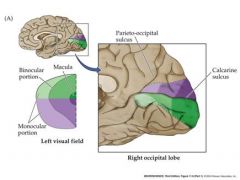
The upper visual field is represented below the calcarine sulcus...lower field above.
left visual field is represented in the right striate cortex |
|
|
|
what do neurons in the primary visual cortex respond to?
experiment |
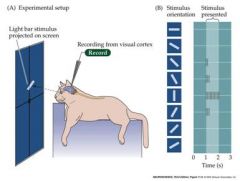
They respond selectively to oriented edges
Cat was anesthetized |
|
|
|
what is the organization like in the striate cortex
|
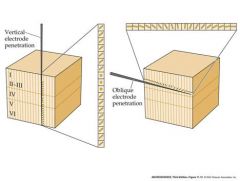
Cells in one vertical column have the same orientation selectivity
Orientation selectivity of columns systematically shifts across v. small regions Of the cortical surface So each little part of the cortex (each part of the visual field) is represented by a complete suite of orientation columns, analyzing line with angles 0 to 180 degrees lateral geniculate axons terminate in cortical layer 4C |
|
|
|
describe how impulses go from the eye to the geniculate nucleus to the striate cortex
|
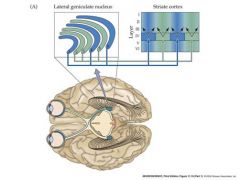
Axons from monocularly-driven
neurons in the Lat. Gen. nucleus terminate on cell bodies of neurons in layer IV of the cortex. The layer IV cortical neurons then send laterally branching axons to synapse on cells in other layers of adjacent cortical areas - these neurons in adjacent areas are then driven by input from both eyes |
|
|
|
draw the picture of the parieto-occipital sulcus and the calcarine sulcus
|
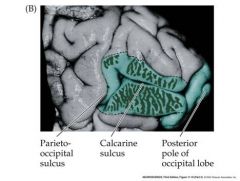
|
|
|
|
talk about binocular disparities
|
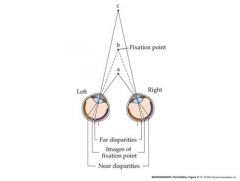
Some binocularly driven neurons in the cortex are maximally stimulated by a particular
Binocular disparity. These cells can signal the distance of an object These cells help to complete the the picture because there are normal disparities...at very large disparities double vision can occur. |
|
|
|
Talk about M and P cells
|
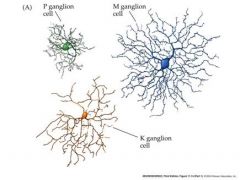
M ganglion cells
Transient response Response not dependent on color Center and surround driven by all cone types P ganglion cells Sustained response Color opponent center surround receptor fields. e.g Red ON center. green OFF surround red light produces a maximal ON response in the center green light produces a maximal OFF response in the surround The center and surround must therefore be driven by different classes of cones Detect color contrast – e,g, a red dot on a green background produces little response, but a red dot on a red background produces a larger response besides layers in the lateral geniculate being specific from each eye...they are also specific for cell size...cells still go to layer 4c in the cortex P ganglion cells send axons to the parvocellular layers of the LGN ...M ganglion cells to the magnocellular layers of the LGN. |
|
|
|
what is the surround created by
|
lateral connections
|
|
|
|
Draw the magno and parvocellular stream
|
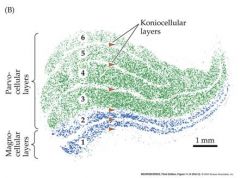
|
|
|
|
talk about extrastriate cortex in monkeys
|
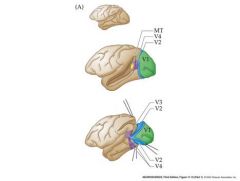
V1 is the primary (striate visual cortex)
To which LGN neurons project Cells in V1 columns send axons To regions V2 etc. Each area (V1, V2, Mt etc) Contains a separate map of the contralateral Visual field Each extrastriate area processes (“contains cells that are especially sensitive to”) a particular set of stimulus features. e.g. MT neurons are maximally stimulated by an edge moving in a particular direction in part of the visual field - Motion sensors. V4 respond selectively to stimulus color without regard to its direction of movement. Lesion, electrical recording and functional imaging studies indicate that information is transmitted between cells in each area by axonal connections along two distinct visual pathways through the extrastriate areas. The ventral (object recognition) pathway V1 > V2 > V4 -color sensitive The dorsal (spatial vision) pathway V1>V2>MT motion sensitive |
|
|
|
draw the two pathways from the visual cortex
|
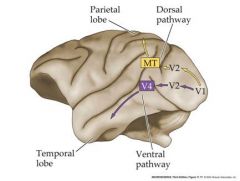
|
|
|
|
what is the binding problem
|
But how is the independent information analyzed in these pathways combined to create a conscious perception of a unified object with color, form and motion?
|
|
|
|
how is the epithelium in the eye arranged
|
It is actually at the back of the eye; thus, the photoreceptors are actually covered by the neurons. The only point this doesn't occur at is at the fovea. The epithelium helps to absorb any light that passes through.
|
|
|
|
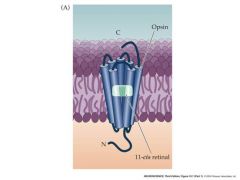
what is rhodopsin composed of?
|
it is composed of opsin and retinal
retinal is a vitamin A derivateve that is the light absorbing portion of the pigment. when activated by light, the two molecules stop binding |
|
|
|
what are sound waves
|
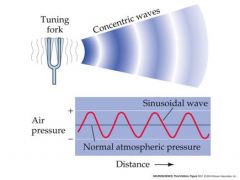
pressure waves of air generated by vibrating air molecules
Sounds are pressure waves of air generated by vibrating air molecules A sound wave has a waveform, phase, amplitude and frequency We can detect sounds having a frequency range of approx 20Hz to 20kHz frequency Complex sounds can be thought of as being the sum of a large number of sine wave components, each having a different frequency amplitude and relative phase (Fourier analysis). So the analysis of the auditory system has often proceeded by first analysing its response to sinusoidal sound stimuli. Amplitude is the max displacement Frequency is the number of full oscillations per second |
|
|
|
talk about phase difference and time delay
|
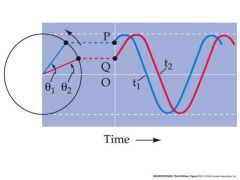
The phase difference (q2 - q1) translates into a time delay, t2 – t1
|
|
|
|
describe the human ear (don't worry about the inner ear)
|
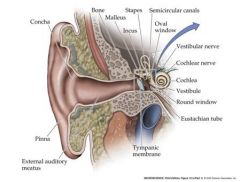
External ear boosts sound frequency 30-100 fold
Eustacian tube is normally collapsed and is attached to the pharynx....when the pressure needs to be balanced, it is opened. oval window and round window are small membranous disks |
|
|
|
how does the middle ear work?
|
The bones of the middle ear act as “impedance matching devices”. They act as a
lever with a high mechanical advantage which efficiently transfers airborne (low impedance) vibrations in the atmosphere to fluid (high impedance) vibrations of the inner ear. Movements of the tympanic membrane due to airborne vibration are focused onto Movements of the oval window and the cochlea fluids beyond |
|
|
|
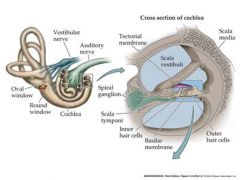
draw cross section of cochlea
|
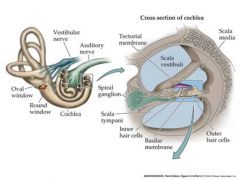
vestibuli is the place where the wave enters....attached to the oval window
Sound leaves out of the scala tympani back into the middle ear. |
|
|
|
Draw the basilar membrane and surrounding structures
|
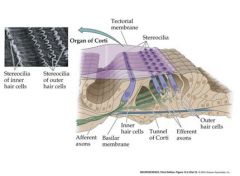
fluid waves push on the flexible membrane of the cochlear duct causing it to transmit a signal
inner hair cells are involved in afferent signals outer hair cells are involved in efferent signals |
|
|
|
What does the basilar membrane of the cochlea contain?
|
sensory receptor hair cells
|
|
|
|
Draw a traveling wave in a cochlea that is uncolied
|
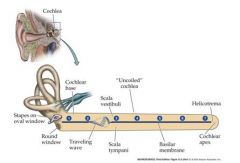
Sound sets up vibration of the oval window, setting up
Traveling waves of basilar membrane vibration that move the down the cochlea helicotrema: place where SV and ST contact. wave goes in the top path first and then through the bottom path |
|
|
|
explain vibration amplitudes in the cochlea
|

Each part of the cochlea
Is “tuned to a separate Frequency sound The amplitude of vibration is greatest in each region at a particular frequency Regions closest to the Stapes are most sensitive To higher frequency Sounds Regions farthest from the Stapes are most sensitive to lower frequency sounds |
|
|
|
draw a picture of shear force
|
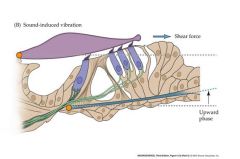
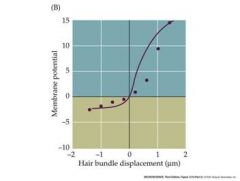
pushing towards the tall stereocilium creates a depolarization....opposite creates a hyperpolarization
Orange points are pivot points Moving basilar membrane causes tectorial membrane to move movement causes stereocilium to move |
|
|
|
draw mechanoelectrical transduction mediated by hair cells
|
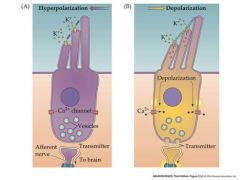
|
|
|
|
scheme for the displacement direction of hair cells and receptor potential
|
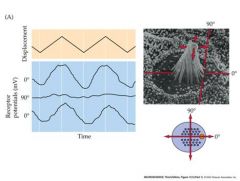
Moving at the 90 degree angle does not do anything.
|
|
|
|
besides having to move in one or the other direction, what else is special about vestibular hair cell?
|
Response is asymmetrical – depolarizing direction
Is more effective |
|
|
|
how does the frequency of a wave influence the depolarization of a hair cell
|

At low frequencies
hair cell Vm follows the vibration of sound, But at high frequencies Only a smooth depolarization persists Asymmetry occurs because of the fact that the response is asymmetrical and depolarizing direction is more effective |
|
|
|
talk about the response properties of auditory nerve fibers
|

A random sampling
Of electrical recordings from single auditory nerve Axons (fibers) reveals that Each axon is tuned (most sensitive to) a particular range of frequencies of sound) lowest peak for each curve is the characteristic frequency |
|
|
|
response properties of auditory nerve fibers all on one page
|
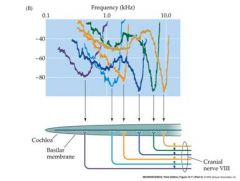
The tuning results
from the fact that Each axon is connected to a hair cell at a different place along The basilar Membrane of the Cochlea. This tuning-by-position Is the only way that The brain can discriminate The frequency (pitch) Of high-frequency sounds (>3 kHz) An axon has to be perceptually “labeled” According to the position of its origin along the Basilar membrane |
|
|
|
what happens above 3kHz in the auditory system?
|
Below 3 kHz, the hair cell membrane potential can “track” the individual peaks of
Pressure during a sound wave (phase-locking), so that the time intervals between action potentials produced by the auditory nerve axon can in theory be used by the brain to determine the frequency of the sound after this the membrane potential is basically stationary |
|
|
|
tonotopic organization
|
neurons are arrayed in brain nuclei and cortex in order of their most-sensitive (characteristic) frequencies
|
|
|
|
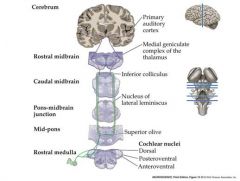
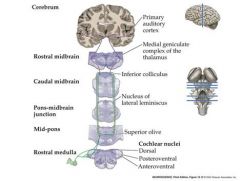
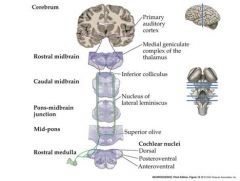
Transmission of information from the auditory nerve sensory neurons to the brain.
|
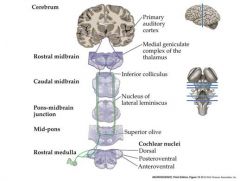
Parallel organization of multiple ascending pathways
Auditory nerve enters the brainstem Branches to innervate 3 subdivisions of the cochlear nucleus Cochlear nucleus neurons project axons to terminate in the ipsi-and contralateral superior olive in the pons and the inferior colliculus in the mid-brain. Inferior colliculus neurons project axons to terminate in the medial geniculae nuceus in the THALAMUS MGN Thalamus neurons send axons to the primary auditory cortex in the temporal lobe |
|
|
|
talk about stereocilia and nerve interaction
|
one nerve branches to many stereocilia, but one stereocilia is only attached to one nerve
|
|
|
|
Processing of sound information at lower brain centers:
|
Role of the medial and lateral superior olives (MSO and LSO) in integrating information from the two ears and localizing the source of a sound
MSO – uses interaural arrival time differences to calculate the direction from which sound is arriving LSO uses interaural intensity differences |
|
|
|
How does the MSO compute the location of a sound
|
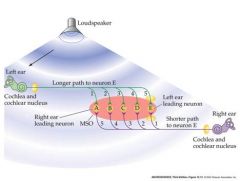
Sound arrives first to left ear...but arrival at neuron E is delayed by a longer axon...path to E is faster from the right ear...sound arrives later though
So, due to the coincident release of NT, activity in E is greatest in response to a sound coming from the left. Neuron A is most sensitive to sound from the right etc The neurons A – E of the MSO act as co-incidence detectors |
|
|
|
How does the LSO work?
|
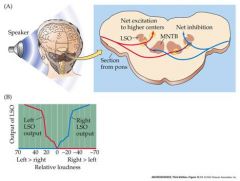
Sound on left side is more intense
– inhibits input from right side -> net Excitation from left hand side sent to higher brain centers indicates sound coming from left Red neuron leading to interneuron to blue neuron is an inhibitory interneuron LSO using intensity differences |
|
|
|
information processing in the inferior colliculus
|
Neurons with cell bodies in the Inferior Colliculus
are arrayed in such a way as to create an auditory space map. neighbouring neurons are preferentially sensitive to sound coming from neighbouring directions. This map is based on the calculated output of the LSO and MSO neurons -Highly developed in nocturnal predators, like barn owls |
|
|
|
information processing in the MGC thalamus
|
Neurons here mediate the detection of specific, complex sound patterns
E.g. echolocating neurons in bat MGC thalamus are sensitive to particular delays between the time of emission of sound and the time at which an echo is received. |
|
|
|
info processing in the auditory cortex
|
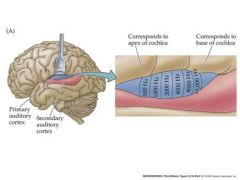
Primary auditory cortex contains a precise tonotopic map – an array of neurons arranged according to their characteristic (best) sound frequencies.
...This is, in effect, a topographic map of the basilar membrane. In addition, cortical neurons can be binaurally stimulated (by both ears) – strips of neurons excited by one ear, but not the other alternate with strips excited by both ears – similar to ocular dominance columns in the visual cortex Cortical neurons are also often particularly sensitive to certain sound combinations that characterize the vocalizations of an animal. |
|
|
|
special part of the auditory cortex
|
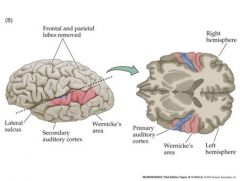
Wernicke's area...important in comprehending speech
|
|
|
|
organization of the human olfactory system
|
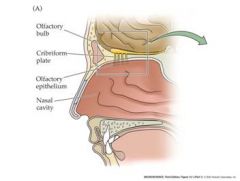
Cribriform plate can become damaged if punched in the nose...it is the white stuff....not the yellow stuff
|
|
|
|
structure and function of the olfactory epithelium
|
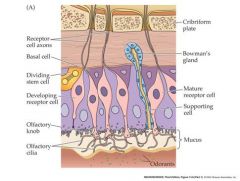
olfactory cilia: inward generator current leads to generator potential
mature receptor cell: generator potential leads to action potential Axons produce the AP |
|
|
|
How often do Olfactory Receptor Neurons renew?
|
every 6-8 weeks
|
|
|
|
Show a stimulation of olfactory cells and the measured current
|
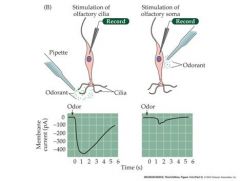
Wholecell patch clamp recordings of generator currents
The cilia are the sensory organelles...contain the transduction machinery |
|
|
|
what is the outer section of a rod or cone actually?
|
it is really a modified cilium
|
|
|
|
olfactory transduction pathway
|
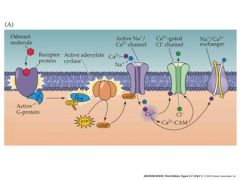
Mucous contains odorant binding proteins that deliver odorant molecules to the cilia
Inward current flows when Odorant is present - generator Current > depolarizes receptor cell Chloride flows out because mucous is low in chloride....net inward current |
|
|
|
what is it when negative ions leave
|
net inward current
|
|
|
|
olfactory receptor molecules
|
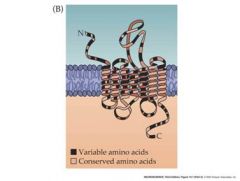
Odorant receptor proteins
are a HUGE Family of related G-protein-linked Receptor proteins Approx 1000 distinct Genes in humans code For odorant receptor proteins! 3-5% of all our genes! (compared to only 4 for opsins.) part of the 7 transmembrane G-pro receptor family |
|
|
|
how many odorant receptor proteins does an olfactory receptor neuron express?
|
1 or at most a few
|
|
|
|
How could you tell different olfactory receptors apart?
|
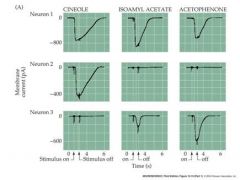
you could use a patch-clamp and see how they responded via their generator current....a downward current is actually a positive voltage
|
|
|
|
What is an important rule about AP's
|
their intensity is always the same, but their frequency can change
|
|
|
|
how does increasing conc of odorant affect AP
|
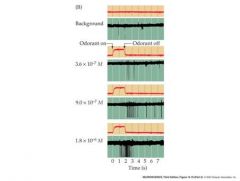
extracellular response of aps in olfactory receptor neuron axons to selected odorants
|
|
|
|
how does each receptor neuron respond?
|
responds with varying sensitivity to a range of odorant molecules...not exclusively to just one
|
|
|
|
if we measure the response of only a single odorant receptor, would it be possible to determine what the receptor was responding to...a high conc odorant or a low conc odorant?
|
no
|
|
|
|
how does the brain compare which odorant is present?
|
compare the output of an array of different odorant receptor neurons and then label the specific pattern of activity in response to a given odor as indicating the presence of that odor.
Presumably stimulation with a particular odorant creates a particular pattern of activity across the olfactory epithelium which is then “mapped” onto neurons in the olfactory bulb – a form of topographical mapping ...brain can't label a given neuron to an odorant ...brain must look at pattern of electrical activity and then it can decipher |
|
|
|
organization of the mammalian olfactory bulb
|
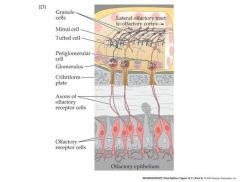
Olfactory receptor neuron (ORN) axons project to
Glomeruli, converging to make synaptic contact with Second-order “mitral cells” In the mouse approx 25,000 ORN’s converge onto 25 mitral cells in each glomerulus – this convergance amplifies the sensitivity for the detection of weak odors Granule cells are localized interneurons that shape the response of the mitral cells don't worry about the tufted cell |
|
|
|
what explains how ORN's expressing a given odorant protein can be so scattered throughout a region of the olfactory epithelium
|
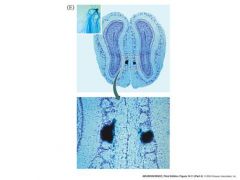
ORN’s that express a
Particular odorant receptor Protein converge onto a subset of glomeruli on each side of a rat’s Olfactory bulb. This convergence explains how ORNs expressing a given odorant peceptor protein can be So scattered Throughout a region of the olfactory epithelium |
|
|
|
how many responses from olfactory receptors can a glomerulus receive?
|
response from thousands of olfactory receptors, but all from the same sub-set....this is the basis of convergence
|
|
|
|
what is special about the pattern of active glomeruli
|
correspond to the pattern of receptor proteins activated by a given odorand....brain can detect previous patterns, it can have an image to oderant
|
|
|
|
draw a pathway of the olfactory tract
|
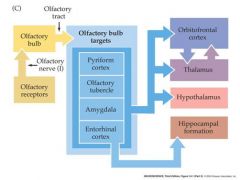
|
|
|
|
organization of the human olfactory system in the brain
|
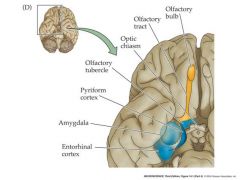
|
|