![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
41 Cards in this Set
- Front
- Back
|
4.1 Inleiding
Leren is het proces waardoor de mens en andere diersoorten kennis over hun omgeving verwerven.
Het geheugen is het reservoir van opgedane kennis dat wordt bewaard.
Terwijl de definitie van 'leren' het actieve en procesmatige aspect van het geheugen beklemtoont, wordt met de definitie van 'geheugen ' vooral |
het passieve en structurele karakter ervan onderstreept. |
|
|
Zelfs zeer primitieve diersoorten beschikken over het vermogen om van hun omgeving te leren en belangrijke inzichten in de theorievorming inzake leren en geheugen zijn gebaseerd op |
onderzoek met dieren met een eenvoudig neuronaal systeem zoals de zeehaas (aplysia).
Cellulaire mechanismen van primitieve leerprocessen als habituatie, sensitisatie en klassieke conditionering werden via onderzoek met ongewervelde dieren ontrafeld. |
|
|
Eerst en vooral willen we inzicht verwerven in de werking van het geheugen. Welke verschillende geheugenfuncties kunnen we onderscheiden ?
|
de cognitieve psychologie die zich vooral vanuit theoretisch en experimenteel oogpunt met het geheugen heeft beziggehouden.
Hoewel de psychologie zich hootdzakelijk vanuit een gedragsobserverende en theoretisch beschouwende manier met het geheugen inliet en aanvankelijk weinig of geen aandacht besteedde aan de biologische aspecten van de geheugenwerking, vormt het cognitief psychologisch model van het menselijk geheugen een belangrijke stap voor een beter begrip van de biologische geheugenwerking. |
|
|
Een tweede vraag heeft uiteraard betrekking op de cerebrale organisatie van geheugen en leren.
We zullen vaststellen dat de psychologische en biologische benaderingen stilaan
|
naar elkaar toegroeiden en dat nieuwe gegevens of hypothesen vanuit de ene discipline |
|
|
Waar de filosofische beschouwingen over het geheugen reeds dateerden vanuit de oudheid en de systematische beschrijving van de geheugen werking zich met de pychologen Ebbinghaus en James aan het einde van de negentiende eeuw situeert, kunnen we de biologische interesse voor het geheugen pas bij het begin van de Eerste Wereldoorlog opmerken.
Gedreven door een lokalisationistische trend trachtte men de cognitieve functies in de hersenen te |
'lokaliseren'.
Lashleys vergeefse queeste naar de plaats waar de geheugeninhouden inde hersenen lagen opgeslagen, maakte komaf met het idee van een lokaliseerbare geheugenfunctie, vergelijkbaar met de door Broca en Wernicke ontdekte taalzones in de linkerhemisfeer. |
|
|
Tegen het midden van de twintigste eeuw geloofden nog slechts weinigen dat geheugen functies in specifieke hersenregio's werden opgebouwd en deze opvatting stond in schril contrast met de lokaliseerbaarheid van vele andere cognitieve functies.
Het geheugen werd beschouwd als |
een algemene eigenschap van de gehele cortex.
Deze sterk globalistische visie nam echter niet weg dat onderzoekers zoals Hebb speculeerden over de biologische grondslag van de geheugenfunctie, zonder evenwel de geheugenwerking in bepaalde hersenregio's te situeren. |
|
|
Een belangrijke doorbraak in bet wetenschappelijk geheugenonderzoek kwam er dankzij de grondige studie van een aantal bijzondere klinische gevallen waarvan de patiënt .... de beroemste is |
H.M. wellicht de meest beroemde is.
Een goedgedocumenteerde neurochirurgische ingreep veroorzaakte bij deze man een onverwacht en dramatisch geheugendeficit dat niet alleen de anatomische basis van een specifieke geheugenfunctie blootlegde, maar tevens een belangrijke stimulans betekende voor de cognitieve theorievorming, en de psychologische en biologische benaderingswijzen dichter tot elkaar bracht. |
|
|
Grondige analyse van geheugendeficits bij H.M. en andere hersenbeschadigde patiënten en bij experimenteel proefdieronderzoek toonde aan dat het geheugen weliswaar in verschillende delen van onze hersenen wordt opgebouwd, maar ..... |
dat bepaalde hersenregio's belangrijker zijn voor de opbouw van bepaalde geheugenfuncties dan voor andere.
Recent neurofysiologisch en biochemisch onderzoek steunt bovendien de eerdere inzichten van Hebb met ontdekkingen als synaptische effectiviteit en langeterrnijnpotentiatie.
|
|
|
Los van elke neuroanatomische context heeft het cognitief psychologisch geheugenonderzoek het geheugen traditioneel voorgesteld al een reeks van bewaarplaatsen die de informatie voor een bepaalde tijd kunnen vasthouden en die met elkaar in verbinding staan.
Naast de bewaarplaatsen onderscheidt men |
een aantal geheugenprocessen die het systeem toelaten de informatie in en tussen de geheugenbewaarplaatsen te manipuleren.
Klassiek wordt het cognitief geheugenmodel voorgesteld als een stroomdiagram met een aantal hokjes die de verschillende geheugen bewaarplaatsen voorstellen en die door middel van pijlen met elkaar verbonden zijn om de interacties tussen de geheugencom ponenten uit te beelden.
Het model vertegenwoordigt een 'standaard ', een hypothese van het gemiddeld normaal geheugenfungeren. Een 'normaal' gebeugenfungeren veronder stelt een correct functioneren van de bewaarplaatsen en van de geheugenprocessen die er plaatsvinden. Het meest bekende geheugenmodel werd geïntroduceerd in de jaren zestig (Atkinson & Shiffrin, 1968 ; Waugh & Norman, 1965) en nadien verder uitgewerkt. Volgens dit model wordt het geheugen verdeeld in drie structurele componenten met specifieke kenmerken |
|
|
Het model vertegenwoordigt een 'standaard ', een hypothese van het gemiddeld normaal geheugenfungeren.
Een 'normaal' gebeugenfungeren veronderstelt een correct functioneren van de bewaarplaatsen en van de geheugenprocessen die er plaatsvinden.
Het meest bekende geheugenmodel werd geïntroduceerd in de jaren zestig (Atkinson & Shiffrin, 1968 ; Waugh & Norman, 1965) en nadien verder uitgewerkt. Volgens dit model wordt het geheugen verdeeld in drie structurele componenten met specifieke kenmerken:
De onderscheiden geheugenbewaarplaatsen zijn: |
• het zintuiglijk geheugen, |
|
|
De onderscheiden geheugenbewaarplaatsen zijn
In deze compartimenten kan informatie worden vastgehouden.
Elke bewaarplaats wordt gekenmerkt door |
een specifieke functie (de rol in de algehele geheugenwerking), een capaciteit (de hoeveelheid informatie die de bewaarplaats op elk ogenblik kan bevatten) en de duur waarmee een stimulus in het geheugencompartiment kan worden vastgehouden.
Naast de verschillende opslagplaatsen specificeert het model een set van controleprocessen zoals patroonherkenning, herhalen , inprenten en ophalen die de interactie tussen de verschillende geheugen bewaarplaatsen weergeven.
|
|
|
Naast de verschillende opslagplaatsen specificeert het model een set van controleprocessen zoals |
-patroonherkenning, -herhalen , -inprenten en ophalen
die de interactie tussen de verschillende geheugen bewaarplaatsen weergeven. |
|
|
Een cognitief-psychologisch model van het geheugen: |
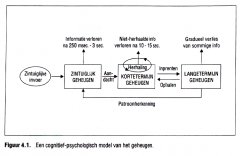
|
|
|
De van de buitenwereld afkomstige prikkels worden via onze zintuigen in een voor de hersenen begrijpbare code omgezet en opgevangen in het zintuiglijk geheugen.
Het zintuiglijk geheugen is een bewaarplaats met een relatief grote capaciteit, maar van een extreem korte duur. Nog merkwaardiger is dat |
wij ons helemaal niet van het bestaan van deze informatiebewaarplaats bewust zijn en dat zij enkel door experimenteel onderzoek kan worden aangetoond.
Hoewel men aanneemt dat er een zintuiglijk geheugen voor elke zintuiglijke modaliteit betaat, werd enkel het visueel en auditief zintuiglijk geheugen beter onderzocht. |
|
|
George Sperling (1960) toonde het bestaan aan van een zeer accurate, maar tevens zeer kortstondige vorm van visuele bewaring, het |
visueel zintuiglijk of iconisch geheugen.
Hij toonde zijn proefpersonen een matrix van drie rijen van elk vier letters gedurende 50 milliseconden (een twintigste van een seconde), een taak die moeilijker is dan lezen met behulp van een bliksemflits. Nadat de twaalf letters van het scherm waren verdwenen, vroeg Sperling zijn proefpersonen zoveel mogelijk items op te zeggen.
Bij deze 'globaal antwoordprocedure' konden de subjecten gemiddeld slechts vier tot vijf van de twaalf letters correct opnoemen .
|
|
|
Hoewel men kan opperen dat de wel bijzonder korte aanbie dingstijd de proefpersonen niet in staat stelde om alle letters te zien, laat staan ze te onthouden, bleek |
dit niet de prestatiebeperkende factor te zijn .
Ook wanneer Sperling de aanbiedingstijd opdreef, bleven de resultaten ongewijzigd. De limiet van het aantal gerapporteerde letters werd dus niet verklaard door de korte aanbiedi ngstijd. Verhoging van de aanbiedingstijd zou dan immers de prestatie hebben moeten verbeteren.
|
|
|
Als de prestatielimiet van de proefpersonen (vier tot vijf items) niet werd veroorzaakt door een beperking in de waarneming (variatie van de aanbiedingstijd toont geen verschillen), dan moet |
het geheugen de limiterende factor zijn, in het bijzonder het aantal items die lang genoeg en simultaan in het kortetermijngeheugen kunnen worden gehouden.
Onder normale omstandigheden 'onthouden' vrijwilligers van een ultrakort aangeboden 3x4- matrix dus een vier- tot vijftal items.
|
|
|
Naast de 'globaal-antwoordprocedure ' ontwikkelde Sperling een 'partieel-antwoordprocedure'.
Tijdens dit experiment, kregen de proefpersonen opnieuw een 3x4-lettermatrix gedurende 50 milliseconden te zien. Onmiddellijk na het verdwijnen van de matrix hoorde het subject een toon van hoge, gemiddelde of lage frequentie. Deze toon was een afgesproken instructie die de proefpersoon vroeg een welbepaalde rij van de matrix op te zeggen. Een hoge toon vroeg naar de letters van de bovenste rij, een gemiddelde toon naar de letters van de middelste rij en een lage toon verwees naar de onderste rij. Onmiddellijk na het horen van de toon probeerden de proefpersonen de letters van de gevraagde rij op te zeggen.
De resultaten van dit experiment bleken |
verbluffend .
Ongeacht welke rij de proefpersonen dienden op te zeggen bleek hun score bijna 100% accuraat ! |
|
|
Dit betekent dat op het ogenblik van het geluidssignaal (en dus nadat de zichtbare lettermatrix reeds was verdwenen) de proef personen alle twaalf letters in het geheugen moesten hebben om een dergelijke accurate prestatie te kunnen leveren.
Bij de globaal-antwoordprocedure bleek dat de proefpersonen slechts vier tot vijf letters konden weerhouden !?
Waarom bestaat er een discrepantie tussen departieel- en de globaal-antwoordprocedure ?
Waarom hebben de proefpersonen een kortetermijngeheugenspan van vier tot vijf letters terwijl ze er in feite twaalf ku nnen onthouden ?
Als het kortetermijngeheugen de eerste vorm van bewaring zou zijn, kunnen we door de limiet van de normale geheugenspan deze resultaten niet verklaren. Om de vraag te beantwoorden dienen we |
een variant van de partieel-antwoordprocedure te onderzoeken.
In de zojuist beschreven procedure volgde de toon vrijwel onmiddellijk na de aanbieding van de letters.
|
|
|
Maar wat zou er gebeuren als het interval letters-toon vergroot ?
Wanneer Sperling het interval tussen het aanbieden van de matrix en de opdracht (geluidstoon) stelselmatig vergrootte tot pakweg één seconde, maakten de subjecten al vlug steeds meer en meer |
fouten tot hun prestaties identiek waren met die van de globaal-antwoordprocedure en er slechts vier tot vijf letters konden worden opgezegd.
|
|
|
De beschikbare informatie onmiddellijk na de visuele aanbieding van de stimulus is dus groter dan wat daar één seconde later van overblijft.
Deze experimenten suggereren dat |
een originele en welhaast fotografische voorstelling van de stimulus in het geheugen wordt opgenomen die evenwel vlug uitdooft zodat met het voortschrijden van de tijd steeds minder informatie over deze stimulus in het geheugen aanwezig is.
Sperling toonde het bestaan van een zeer accurate maar tevens zeer kortston dige vorm van visuele of iconische bewaring naast het gekende kortetermijngeheugen. De term icon verwijst naar het interne fotoachtige beeld van een niet meer aanwezige visuele stimulus. Door manipulatie van bet stimulus-tooninterval kon de duur van het iconisch geheugen op 250 msec. worden geschat.
De term iconische bewaring of iconisch geheugen verwijst naar de residuele neurologische activiteit in de visuele areas van de cortex wan neer de externe, reële visuele stimulus niet meer aanwezig is. |
|
|
Sperling toonde het bestaan van |
een zeer accurate maar tevens zeer kortstondige vorm van visuele of iconische bewaring naast het gekende kortetermijngeheugen. De term icon verwijst naar het interne fotoachtige beeld van een niet meer aanwezige visuele stimulus. Door manipulatie van bet stimulus-tooninterval kon de duur van het iconisch geheugen op 250 msec. worden geschat.
De term iconische bewaring of iconisch geheugen verwijst naar de residuele neurologische activiteit in de visuele areas van de cortex wan neer de externe, reële visuele stimulus niet meer aanwezig is. |
|
|
De term iconische bewaring of iconisch geheugen verwijst naar |
de residuele neurologische activiteit in de visuele areas van de cortex wan neer de externe, reële visuele stimulus niet meer aanwezig is. |
|
|
Meer direct bewijs dat het iconisch geheugen informatie in zijn originele zintuiglijke vorm bewaart, werd geleverd door Eriksen en Collins (1967).
Zij ontwierpen paren van stippenpatronen die, wanneer ze afzonderlijk werden bekeken, een schijnbaar willekeurig stippenpatroon lieten zien.
Wanneer de gepaarde afbeeldingen echter op elkaar werden geprojecteerd, vormden ze samen een drieletterwoord.
Proefpersonen bleken in staat om |
in het simultaan aangeboden stippenpatroonpaar het drieletterwoord te herkennen.
Ze bleken ook in staat het drieletterwoord te herkennen wanneer de afzonderlijke stippenpatronen met een interval van minder dan 300 milliseconden na elkaar werden getoond.
|
|
|
Dankzij het visueel zintuiglijk geheugen versmolt het icon van de eerste afbeelding met de |
aanbieding van de tweede afbeelding zodat beide stippenpatronen als één geheel werden waargenomen en het drieletterwoord herkenbaar werd.
Wanneer het aanbiedings interval tussen beide aanbiedingen werd vergroot, zagen de proefpersonen nog slechts afzonderlijke stippenpatronen en konden ze het drieletterwoord niet meer waarnemen.
Misschien nog meer dan in het experiment van Sperling wordt hier de bewaringsfactor van het iconisch geheugen geïllustreerd. |
|
|
Wanneer het aanbiedings interval tussen beide aanbiedingen werd vergroot, zagen de proefpersonen nog slechts |
afzonderlijke stippenpatronen en konden ze het drieletterwoord niet meer waarnemen.
Misschien nog meer dan in het experiment van Sperling wordt hier de bewaringsfactor van het iconisch geheugen geïllustreerd. |
|
|
4.4 De functie van het zintuiglijk geheugen
De informatie verblijft slechts een korte periode in het zintuiglijk geheugen . Hoe langer zij er verblijft, hoe zwakker zij wordt, tot ze geheel uitdooft. Wat is de functie van deze getrouwe maar zeer kortstondige bewaring ?
|
een vrij primitieve geheugenvorm die het materiaal in een precategorische vorm bewaart.
Met precategorisch bedoelt men de bewaring alvorens de stimulus door patroon herkenning tot een bepaalde categorie wordt toegekend.
|
|
|
Een tweede variant van Sperlings partieel-antwoordprocedure verduidelijkte het precategorische karakter van de zintuiglijke bewaring.
In dit experiment bestond de matrix uit willekeurig gerangschikte letters én cijfers. Het geluidssignaal vroeg dit keer niet naar een bepaalde rij, maar naar een bepaalde categorie (cijfers of letters). De hypothese stelt dat in een precategorisch icon de proefpersonen geen onderscheid kunnen maken tussen beide soorten stimuli omdat ze slechts als visuele patronen worden waargenomen en nog niet als een cijfer of letter worden herkend .
De resultaten van het experiment |
bevestigen de hypothese. De prestatie van de proefpersonen bleek identiek met die van de globaal-antwoordprocedure. |
|
|
De voornaamste functie van het zintuiglijk geheugen bestaat erin |
de ruwe informatie langgenoeg vast te houden zodat de hogere perceptuele functies de informatie voldoende kunnen verwerken en vergelijken met de reeds aanwezige kennis.
Dit waarnemings- en herkennigsproces vraagt echter tijd en zou niet naar behoren kunnen werken indien onze waarneming zich tot de reële duur van de stimuli zou beperken. Terwïl het materiaal in het iconisch geheugen wordt vastgehouden , wordt het door middel van patroonherkenning geanalyseerd en gecategoriseerd.
Patroon herkenning definiëren we als de vergelijking van nieuwe ·zintuiglijke informatie met de reeds aanwezige informatie in het permanente of langetermijngeheugen. |
|
|
Dit waarnemings- en herkennigsproces vraagt echter tijd en zou niet naar behoren kunnen werken indien onze waarneming zich tot de reële duur van de stimuli zou beperken. Terwïl het materiaal in het iconisch geheugen wordt vastgehouden , wordt het door middel van patroonherkenning geanalyseerd en gecategoriseerd.
Patroon herkenning definiëren we als |
de vergelijking van nieuwe zintuiglijke informatie met de reeds aanwezige informatie in het permanente of langetermijngeheugen. |
|
|
Het doel van de patroonherkenning is
|
de voor het systeem relatief nutteloze infomtatie zoals visuele vormen of spraakpatronen te vertalen in iets dat wel een betekenis heeft.
Patroonherkenning is een complex proces van beperkte capaciteit waarbij contact wordt gemaakt tussen de in het zintuiglijk geheugen aanwezige informatie en de reeds verworven kennis.
Tijdens dit proces worden de voor het individu relevante prikkels, dat wil zeggen die prikkels waarvan de zintuiglijke aspecten enigszins overeen komen met betekenisvolle concepten, 'herkend ' en belanden ze in een tweede geheugenopslagplaats, het kortetermijngeheugen.
Wanneer de minimale verwerkingstijd van een visuele stimulus op ongeveer 200 milliseconden wordt geschat, komt dit goed overeen met de levensduur van het icon en de functie die het wordt toebedeeld (Eriksen & Eriksen, 1971). |
|
|
Nu we het bestaan van het iconisch geheugen hebben aangetoond en zijn functie bij de stimulusherkenning hebben gesitueerd, kunnen we ons afvragen hoe het komt dat we vloeiende beelden kunnen waarnemen en geen reeks van op elkaar volgende stilstaande plaatjes ?
De reden is dat
|
het icon sterk wordt beïnvloed door de stimuluscondities zoals het contrast van de stimuli en de helderheid van de achtergrond.
Het icon blijkt in het bijzonder gevoelig voor een volgend icon dat het eerste icon als het ware overschrijft.
|
|
|
Dit effect wordt achterwaartse maskering genoemd en vermijdt een opeenstapeling van iconische beelden die de visuele informatie troom zou vervormen.
Het zintuiglijk geheugen voor verbaal materiaal wordt |
akoestisch of echogeheugen genoemd (Neisser, 1967).
Het auditief zintuiglijk geheugen waarvan de duur op 150 msec. tot 3 sec. wordt geraamd, speelt eveneens een belangrijke rol bij de herkenning van zeer korte stimuli.
|
|
|
Zonder een echogeheugen zou het begrijpen van gesproken taal enorm bemoeilijkt worden vermits we enkel de fysiek aanwezige geluiden zouden kunnen horen. Intonatieverschillen in een vragende of bevelende zin zouden de toehoorder ontgaan.
Woordverbeteringen zoals "Nee, niet peer, beer !" hebben slechts bun effect wanneer de 'p' in peer lang genoeg aangehouden kan worden om de vergelijking met 'b' uit beer mogelijk te maken.
Juist omdat de (gesproken) taal een sequentiële orde heeft, is |
een zintuiglijke bewaarplaats voor haar temporele componenten noodzakelijk (Moray, Bates & Barnett, 1965 ; Darwin, Turvey & Crowder, 1972).
En is de duur van het echogebeugen ook langer dan die van het iconisch geheugen dat vooral spatiële informatie zal verwerken. |
|
|
4.5 Het kortetermijngeheugen en de informatiestroom
Nadat de initiële stimuli voldoende lang in het zintuiglijk geheugen werden gehouden opdat patroonherkenning zou kunnen plaatsvinden, worden de herkende patronen door gegeven aan het |
kortetermijngeheugen.
Vermits de patroonherkenning een vergelijkings proces met bekende patronen (uit bet langetermijngeheugen) veronderstelt, komt de informatie die in het kortetermijngeheugen wordt opgenomen, theoretisch gezien uit het langetermijngeheugen en slechts onrechtstreeks uit het zintuiglijk geheugen.
|
|
|
Toch moeten we onderscheid maken tussen de invoer van nieuwe informatie die vanuit de buiten wereld ons kortetermijngeheugen binnendringt en het zich bewust herinneren van voorbije gebeurtenissen of kennis inhouden die vanuit het langetermijngeheugen moeten worden opgehaald.
Beide bronnen dragen bij tot een voortdurende stroom van gedachten, gevoelens en waarnemingen die het kortetermijngeheugen vullen.
Het kortetermijngeheugen is |
een opslagplaats met een beperkte capaciteit, in elk geval beperkter dan die van het zintuiglijk geheugen.
Herinner u dat de proefpersonen van Sperlings lettermatrix ook na langere aanbieding slechts vier tot vijf letters bewust konden onthouden.
|
|
|
Gezien de beperkte capaciteit van het verwerkings- en geheugensysteem is een selectie van informatie noodzakelijk.
Het subject wordt immers voortdurend met informatie overladen en is verplicht een deel van de informatie te negeren en een ander deel te selecteren ter verwerking, herhaling en con olidering.
Enkel die stimuli waarop wij onze aandacht richten, en vaak zijn dat de nieuwe, overwachte of veranderende stimuli, dringen tot ons kortetermijngeheugen door.
Dit neemt niet weg dat de |
totale hoeveelheid infomatie die gedurende minuten of uren doorheen het kortetermijngeheugen stroomt, enorm is.
De meeste informatie wordt echter niet bewust geregistreerd en zal niet in ons geheugen worden opgeslagen. |
|
|
De kortetermijngeheugenbewaarplaats kreeg in de wetenschappelijke literatuur een verwarrende hoeveelheid namen toebedeeld.
Naast de term kortetermijngeheugen werd deze bewaarplaats ook wel werkgeheugen, onmiddellijk geheugen of primair geheugen genoemd.
Soms wordt het kortetermijngeheugen ook vergeleken met het bewustzijn, waarmee men wil aangeven dat |
in dit geheugen de informatie verblijft waarvan wij ons bewust zijn, waar we op bet ogenblik aan denken, waar we mee werken.
Hoewel het vreemd kan lijken het menselijk bewustzijn met een geheugenopslagplaats te vergelijken, mag men niet vergeten dat om een denkproces mogelijk te maken men over informatie moet beschikken waarover deze gedachten zullen gaan en dat de informatie in herinnering gehouden moet worden terwijl men erover nadenkt. |
|
|
Hoewel het vreemd kan lijken het menselijk bewustzijn met een geheugenopslagplaats te vergelijken, mag men niet vergeten dat om een denkproces mogelijk te maken men over |
informatie moet beschikken waarover deze gedachten zullen gaan en dat de informatie in herinnering gehouden moet worden terwijl men erover nadenkt. |
|
|
4.6 De capaciteit van het kortetermijngeheugen
De limiet waaronder een onmiddellijk en perfect nazeggen mogelijk is, wordt de geheugenspan genoemd en beschouwd als de geheugenlimiet van het kortetermijngeheugen.
Hoewel de schatting van de grootte van het kortetermijngeheugen afhankelijk blijkt van de taak waarmee ze wordt gemeten, levert de geheugenspantechniek een capaciteit van om en bij de zeven items op (Miller, 1956). Wij blijken in staat om een lijst van uitspreekbare stimuli in het kortetermijngeheugen te houden door ze steeds |
opnieuw bij onszelf te herhalen.
Na elke herhaling sterft het geheugenspoor uit en zou het volledig verloren gaan mocht het niet door een nieuwe herhaling worden verlevendigd. De capaciteit van het kortetennijngeheugen wordt bijgevolg bepaald door de snelheid waarmee we mentaal van het ene naar het andere item kunnen springen om het te herhalen en door de snelheid waarmee elk geheugen poor uitdooft wanneer het item niet wordt herhaald.
|
|
|
Wanneer we een langere set van stimuli proberen te onthouden, wordt het moeilijk om ze allemaal te overlopen en te vermijden dat de eerste is uitgedoofd alvorens we de reeks opnieuw moeten herhalen.
Toch is dit precies wat men probeert wanneer men een zojuist opgezocht telefoon nummer bij zichzelf herhaalt terwijl men naar het telefoon toestel spurt. Men kan deze prestatie gerust vergelijken met het circusnummer waarbij de jongleur een toenemend aantal borden in beweging moet zien te houden om te vermijden dat ze van hun stok tuimelen.
Men weet ook dat deze taak de volledige aandacht vereist en dat een korte afleiding voldoende is opdat het telefoonnummer uit zijn geheugenbuffer zou verdwijnen .
Onderzoek toont aan dat indien de items niet worden herhaald , zij na ongeveer |
tien à vijftien seconden zijn uitgedoofd en uit het kortetermijngeheugen ver dwijnen (Peterson & Peterson , 1959). Merk op dat de uitdovingstijd aanzienlijk langer is dan die van het zintuiglijk geheugen . |