![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
74 Cards in this Set
- Front
- Back
|
What is the goal of ENCODE? |
to fully describe the list of functional elements that make up the human genome |
|
|
"wobble" |
non-Watson/Crick base pairs where a nucleotide can pair to a few different mRNA nucleotides |
|
|
Aminoacyl - tRNA synthetase |
1) forms a phosphodiester bond between an amino acid and ATP to get aminoacyl-AMP 2) forms a bond between aminoacyl-AMP and tRNA to get tRNA with amino acid on it |
|
|
Inosine |
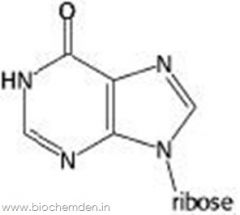
|
|
|
Dihydrouridine |
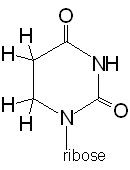
found in the D loop always in the same position |
|
|
Pseudouridine |
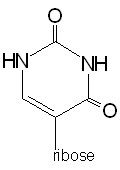
in the TψC loop |
|
|
A site |
"aminoacyl tRNA" site, where individual tRNAs enter the ribosome |
|
|
P site |
"peptidyl site" where growing chain of amino acids come out of the ribosome |
|
|
E site |
the exit site for tRNA |
|
|
IF-3 |
initiation factor in bacteria that binds to the small subunit, blocks the exit site |
|
|
IF-1 |
initiation factor in bacteria that binds near the A site to prevent any tRNA from coming into the A site so only P site is available |
|
|
IF-2 |
Initiation factor in bacteria that interacts with tRNA(fMet) and can't bind to any other tRNA has GTP activity - when large subunit is bound, GTP is hydrolyzed to GDP, IF-2 loses affinity and it ejected |
|
|
tRNA(fMet)
|
initiator tRNA where f stands for formal, has formylated amino group to prevent formation of a chain in front of it, goes into the P site |
|
|
Shine Delgarno Sequence |
sequence in bacterial mRNA, upstream of the start codon, helps recruit the ribosome to the mRNA by aligning the ribosome with the start codon - binds to 16s rRNA when AUG is in the P site |
|
|
riboswitches |
regulatory segment of mRNA that binds a small molecule - flips RNA between 2 structures (can bind Shine Delgarno, surround tRNA and block ribosome, can be removed)
|
|
|
eIF-3 |
eukaryotic initiation factor that binds to 40s subunit and recruits complex with EIF2, tRNA(fmet) and GTP carrying Met also interacts with eIF-4G |
|
|
eIF-4F |
is recruited along with mRNA when Met is added to the 40s subunit, recognizes capped mRNA consists of eIF-4E, -4A, and -4G |
|
|
Kozack sequence |
sequence in eukaryotic mRNA that is recognized by the 18s rRNA, aligns the ribosome with the start codon and helps start translation |
|
|
eIF-4E |
cap binding protein, directs ribosome to the mRNA cap |
|
|
eIF-4A
|
helicase, uses ATP hydrolysis to melt secondary structure of mRNA so the ribosome can scan for the start codon |
|
|
eIF-4G |
the "glue," involved in protein-protein interactions, binds the polyA binding protein that coats the polyA tail of the mRNA -> makes the structure of mRNA circular so translation is more efficient |
|
|
eIF-2 |
a G protein that facilitates binding of Met to the small euk subunit |
|
|
eIF-2B |
guanine exchange protein that puts GTP on eIF-2 and removes GDP, thereby activating it |
|
|
heme controlled repressor |
kinase that phosphorylates the GTP-alpha subunit of eIF2 when the cell is experiencing heme starvation; after eIF2 ads Met to the small subunit and binds eIF-2B, GTP and GDP are exchanged but eIF2 and eIF-2B can't dissociate, this makes eIF2 unavailable so translation halts |
|
|
IRES |
internal ribosome entry site found on viral mRNA, does not depend on cap structure so after the virus stops host translation by decapping its mRNA can still be translated |
|
|
EF-Tu |
bacterial elongation factor that binds aminoacyl tRNA and GTP and has a high affinity for the ribosome; delivers tRNA |
|
|
GAC |
GTPase activating center in bacteria, part of the ribosome in the large subunit that catalyzes hydrolysis of GTP on EF-Tu (which puts it into low affinity so it dissociates) |
|
|
EF-Ts |
GTP exchange factor for EF-Tu |
|
|
EF-G |
binds and promotes movement of tRNA through the bacterial ribosome, also promotes movement of the ribosome down the mRNA - binds to A site so small subunit ratches back and forth counterclockwise, EF-G stabilizes structure -also has G protein activity, when it gets hydrolyzed and changes conformation the ribosome ratchets back to move down the mRNA -mimics EF-Tu when they are bound to tRNA |
|
|
Accomodation |
process by which 2 adenosines in 1st codon position swing up and interact with the minor groove of mRNA, if codon/anticodon pairing is correct, a G in the 2nd position will cause a stacking interaction (no real interactions in 3rd due to wobble) this stabilization is needed to proceed further |
|
|
Paromomycin |
drug that shifts adenosines to "up" position without the right codon/anticodon pairing so misreading occurs |
|
|
Experiment to prove which part of the ribosome is the peptidyl transferase |
Treat 50s subunit with protease so only 23s rRNA is left; it still supports 87% of activity and peptide bond formation |
|
|
Hyrbidized State |
forms after peptide bond occurs; acceptor stem in A site is moved over to the P site, and deacylated tRNA has affinity for the exit site |
|
|
Release Factors |
-recognize stop codons, hydrolyzes the peptide on tRNA to release it - when RF binds it opens up so GGQ can be involved with hydrolysis and other sequence can recognize the codon -mimics the structure of tRNA when it is open -RF1 and RF2 have different conserved sequences and recognize different stop codons |
|
|
GGQ motif |
motif in the release factors that is involved with hydrolysis of the peptide on tRNA, may be involved with G protein activity mimics the 3' end of the animoacyl group of tRNA (CCA) |
|
|
Nonsense mediated decay |
process to get rid of RNA with an accidentally added stop codon -recognized as "bad" if an exon junction complex is downstread of the "stop codon" so the ribosome slows down, RFs bind, proteins build a bridge to the exon junction complex and phosphorylate it and gets sent to processing bodies |
|
|
processing bodies |
densities withing the cytoplasm where RNA is degraded; mRNA gets decapped, deacylated and hydrolyzed down as part of nonsense mediated decay |
|
|
siRNA |
short interfering RNA, protects the prokaryotic cell from viral genome |
|
|
Dicer |
involved in producing siRNAs; binds to 3' end of dsRNA, cuts it (structurally homologous to RNase 3), gives a staggered end and always a specific length also has PAZ domain: 3' overhang binding pocket also cuts pre-miRNA before its tranfered to RISC |
|
|
RISC |
formed from one of the strands cleaved by Dicer and the protein Ago2 Activity is determined by the strand, or "guide sequence" Ago2 has nuclease activity, allowing RISC to cleave complimentary mRNA (which can generate more small RNA and amplify effects) can also engage polyA tail - translational repression |
|
|
miRNA |
small RNA that regulates ~50% of the human genome, targets mRNA at the 3" UTR end for repression or degradation consists of strand cleaved by Dicer from pre-miRNA and transferred to Ago/RISC |
|
|
pri-miRNA |
the origin of miRNA; has lots of stem loop structure, ds stem is cut by Drosha |
|
|
Drosha |
cuts pri-miRNA 11nt up the stem to create pre-miRNA |
|
|
DGCR8 |
recognizes region between ss and ds on pri-miRNA and orients Drosha |
|
|
pre-miRNA |
precursor miRNA with a 2 nt overhang at the 3' end, gets cleaved by Dicer |
|
|
piRNA |
piwi-associated RNA; part of the Ago family and looks like Ago structurally; stops transposon activity by preventing RNA-interferance activity in germ cells |
|
|
GW182 |
bound to Ago2, has affinity for poly-A binding protein can also bring things into processing bodies can bind NOT1 complexes with multiple nucleases that interact with polyA tail, also recruits decapping proteins |
|
|
NOT1 |
binds to GW182 can bind eIF4A1 (helicase) and prevent scanning, or eIF4AII and block movement of subunit (?) |
|
|
Ago2 |
binds the guide strand to make RISC can also engage 4E bound to the mRNA cap, interaction prevents 4G and 4A from binding PAZ domain binds 3' overhang, piwi domain does cleavage and binds GW182 |
|
|
seed region |
nts 2-8 on miRNA guide sequence that must have perfect base pairing with target RNA |
|
|
PAN2/3, CAF1, and CCR4 |
polyA nucleases recruited by GW182 |
|
|
DCP2 |
decapping enzyme recruited by GS182 |
|
|
XRN1 |
exonuclease |
|
|
RITS |
"RNA-induced transcriptional silencing" occurs in the centromere region where genes are repressed until S phase (heterochromatin) Ago1 with guide RNA from centromere targets mRNA as it it made in the nucleus |
|
|
ORI |
origin of replication, consisting of three 13-mers and four 9-mers |
|
|
DnaA |
bind to 9-mers on ORI and opens up the strands, bacterial |
|
|
DnaB |
binds to ORI after DnaA has opened up the strands and stabilizes open complex Has helicase activity, prevents unwinding, bacterial |
|
|
DnaG |
binds to ORI after DnaB, is a primase that makes the RNA primer that polymerase III will use, bacterial |
|
|
primosome |
DnaA, B, and G complex |
|
|
Ago1 |
during RITS, binds guide RNA from centromere and recruits methylase, H3H9 (repression) and attracts Swi 6 (recompaction) inactivates genes in the centromere |
|
|
Dam Sequence |
sequence at Ori (GATC) where the As can get methylated |
|
|
SeqA |
binds tightly to hemimethylated Dam sequence after replication, prevents binding to Ori and shuts off replication temporarily binds the rest of DNA as it gets replicated and hemimethylated (which allows Ori to get fully re-methylated) |
|
|
single stranded DNA binding protein |
on lagging strand, protects it |
|
|
DNA gyrase |
DNA unwinding |
|
|
B clamp |
encloses DNA during replication, put on by the clamp loader, interacts with DNA Pol III during replication and dissociates from it when Pol III reaches end of Okazaki fragment, left behind on DNA recruits Pol I to remove RNA primer from Okazaki fragment and replace it with DNA, recruits DNA ligase to fix nick between fragments |
|
|
Bgl II |
restriction site used to cut fragments of DNA for Okazaki frgament experiment - end up with large fragment from primer to Bgl II that can't fit through the gel and a small fragment from Bgl II to primer |
|
|
clamp loader |
y-complex, an AAA+ ATPase, opens up 1 of 2 dimer interfaces on B-clamp and guides it onto 3' end of primer, then senses duplex and the junction and dissociates delta subunit is in excess and exists independently of the loader, can trigger release of B clamp from DNA |
|
|
pol-alpha |
in euks, puts down primer that is combination of RNA/DNA |
|
|
ORC |
euk origin of replication complex, stays on DNA throughout cell cycle, binds cdc6, cdt1, and mcm2-7 during G1 phase |
|
|
licensing complex |
euk pre-replicating complex composed of ORC, cdc6, cdt1, and mcm2-7; in S phase kinase activities are activated to inactivate proteins so so pre-replication complex doesn't start replication again |
|
|
cdc45 and GINS |
cause strand separation in euk replication, recruited by the replicative complex |
|
|
pol I |
has 5'-3' exonuclease activity, can removed RNA primer and put down DNA |
|
|
DNase I hypersensitive sites
|
genomic regions that are accessible to enzymatic cleavage as a result of the displacement of nucleosomes
|
|
|
replisome |
DNA pol III bound to the primosome (Dna A, B, and G) |