![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
38 Cards in this Set
- Front
- Back
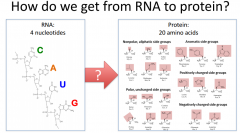
How do we get from RNA to Protein? |
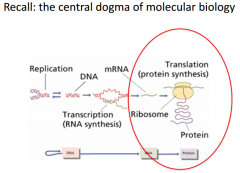
We use tRNA (our adapter molecule which helps communicate and translate between proteins. Occurs during translation (protein synthesis) |
|
|
Describe the basic structure of a tRNA |
It has a region where it can base pair with 3 other nucleotides on an mRNA codon. However, we know that the tRNA has a much more complicated structure. It's considered to have a cloverleaf structure. tRNA goes through a series of processing that will involve the addition of non-canonical nucleotides, which means that a tRNA molecule undergoes a lot of modifications. The "business" end of the tRNA is called the anti-codon loop. |
|
|
Anti-Codon Loop |
The part of the tRNA that makes contact and hydrogen bonds with the base pairs of the codon on the mRNA. There is going to be a reverse orientation when we describe codons and anti-codons. The 5' end of the anti-codon is going to be base pairing with the 3' end of the codon and vice versa. |
|

|
A. On the mRNA, the sequence would be UGA because it's the same as the DNA but replaced T with U. If you write UGA in a 5' to 3' direction, the anti-codon ACU would be able to pair with it. |
|
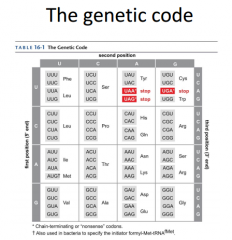
|
These are the different anti-codons that will code for our mRNA codon. Amino acids have more than 1 codon that codes for them. This means it's degenerate, because they have more than 1. |
|
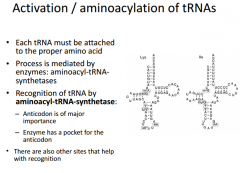
|
We have to activate out tRNAs in order to have them participate in translation. tRNA activation is called aminoacylation of tRNA. This means we connect an amino acid to the top end of the tRNA. Each tRNA will be specific for 1 amino acid. We have tRNAs with different anti-codons but the same amino acid linked to them. |
|
|
What is the enzyme that carries out this process? |
Aminoacyl-tRNA synthetase. There's going to be 1 of these enzymes for each amino acid because they are going to have a specific binding pocket that works for only 1 amino acid |
|
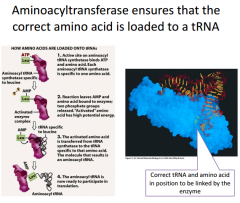
Looking at the reaction up close |
The purple blob is the eznzyme. It will bind one amino acid (the one it's specific for) and ATP. In the active site for this enzyme, we're going to hydrolyze ATP and keep the adenisine monophosphate linked on to the amino acid. This is now a very high energy amino acid that has an AMP molecule covalently linked to it. So we'll have a pyrophosphate as our leaving group. Step 2: Synthetase will bind a tRNA molecule. When it does that, it will transfer the amino acid onto the tRNA. |
|
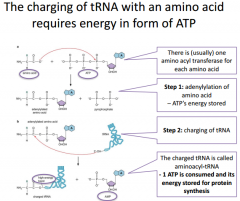
An even closer look at reaction |
Our first reaction is going to be transfering ATP energy onto the amino acid, making it a higher energy molecule. Then we're going to transfer this onto the tRNA. On one end of the tRNA we have a 3' hydroxyl and use this to covalently link an amino acid to it. The AMP is going to be the leaving group. The bond that links the tRNA to the amino acid is considered a high energy bond, and this makes it so that we don't actually have to consume ATP energy during peptide bond formation during translation because the energy is already stored in the bond that's in the activated or charged tRNA. |
|

Describe experiment (Nurimberg Experiment) No CQ. Answer is A. |
We've taken everything out of the cell that we need in order to translate a protein. We work this these materials outside the cell. We have a cell extract (contains translation machinery). In this experiment, they took all 20 aa and varied one by one which aa was radioactive. They then attempted to translate an mRNA molecule into protein. If a protein is made and it's radioactive we can capture that on a membrane and detect it. The only way the protein will be radioactive is if we added in the correct radioactive tRNA containing the aa for which our mRNA molecule encodes. We had an RNA sequence that was just a string of Uracils. They performed these many reactions where they varied aa and radioactivity. They tried to see if a radioactive protein was made. The only time they saw a radioactive protein made was when they had tRNA covalently linked to Phenylalynine. Phe is encoded by UUU. This is how they were able to first crack the genetic code. |
|

|
C. If you just have the same nucleic acids (a bunch of Us), you're not going to be able to tell if it was the first 3 Us that encoded the aa, or if it was ther 2, 3 and 4th etc |
|
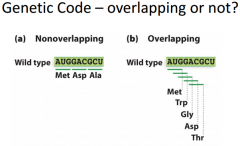
To help visualize |
a) We have a frame we have to follow B) The A, the U or G etc. could be the first position, and we could encode 5 amino acids which isn't right Genetic code must be non-overlapping |
|

|
. |
|
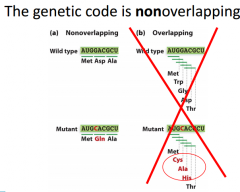
|
If we have 1 base pair mutation that only leads to 1 aa mutation, this is a clue the genetic code is nonoverlapping. For overlapping, If we were to mutate this base here, we would be changing 3 different aa positions. |
|

|
There was a problem with actually having to analyze the addition codons. Once we were able to synthesize molecules with different nucleotides, we were able to identify other molecules and find the other codons. |
|

STOP and START Codons |
We have 3 stop codons. These codons don't code for a particular aa, they just code for a stop (termination). Translation termination factors will recognize these codons. We also have a start codons. We will always start with a Met, but that doesn't mean we won't have these Mets throughout the sequence. Their will be a difference when it's considered a START AUG and a internal AUG. Start AUG will bind initiator MET tRNA, regular AUG will bind regular tRNA that will code of regular MET on it. |
|
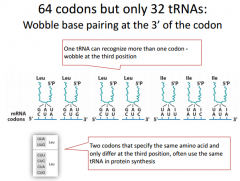
|
These are all of the mRNA codons that code for Leu. The mRNA is attached to the anti-codon. We have 7 different possibilities here, and we will have different ways to create multiple codons for an mRNA codon. |
|
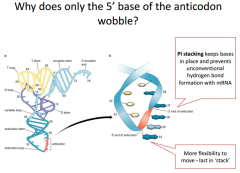
|
The wobble position is the 1st position of the tRNA anti-codon reading from 5' to 3'. If we looked at it from the 3' to 5' direction it would be the 3rd position. In the 1st position (red) we have a lot more room for flexibility for what that position could be because it's not as sterically hindered as the other 2 positions (they follow bonding rules and match canonically). This 1st position doesn't need to follow the rules and doesn't need to bind canonically. We have a little bit of wobble room and can base pair with different nucleic acids. Wobble allows one tRNA to recognize 2 different codons. |
|
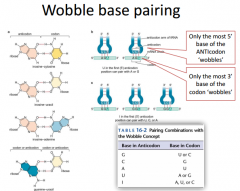
Wobble Table |
The wobble table shows which base in the anti-codon is going to base pair with one of the codons. We have this unusual base called Inicine. Inicine can make a base pair with A, U or C. These are the non-canonical base pair rules. |
|

|
3. For the 1st one, the first two base pairs are the same. For the 2nd one, the first two base pairs are also the same. |
|

To help explain answer |
|
|
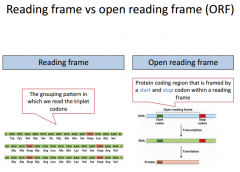
|
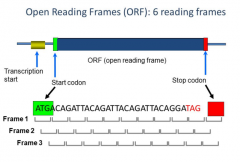
RF can be read by every 3 nucleotides starting from a particular position. The ORF is from right before the start codon to right before the STOP codon. The ORF is all of the codons that are actually translated into an amino acid. We do not include the STOP codon because it doesn't encode for any aa. We have several different reading frames we could encode from here. |
|

|
We have 6 different reading frames. |
|
|
There are 6 different ways we could read the frame, although it really is only supposed to be 1. |
|

|
C. |
|
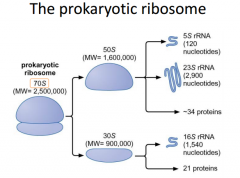
Need to know prokaryotic subunits |
The ribosome is the translation machinery of a cell. It consists of 2 main subunits. Within these subunits, we have a number of different proteins and RNA molecules. We call the 2 ribosomal subunits the 70s unit. Seperately, they are called the 50s and the 30s. The S stands for Sedberg Units. In the large subunit, we have different RNA molecules. The 5s rRNA (ribosomal RNA) and the 23s rRNA (ribosomal RNA). The smaller subunit has the 16s rRNA. |
|
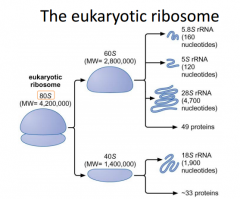
DONT need to memorize eukaryotic factors |
Large, more components. |
|

|
C. RNA with catalytic activity. Ribozymes are components of the ribosomal RNAs that are part of the ribosome. These ribosomal ends are the important part. The protein ends are just their to orient the RNAs in the correct place to interact with mRNA/tRNA molecules to form the peptide bonds. |
|

|
B. Prokaryotes will have transcription/translation co-occurying at the same time. So RNA is synthesized in the cytoplasm while the ribosomes also translate the RNA. (Euk must splice introns and transport etc). |
|
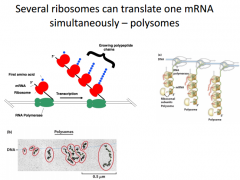
|
You can get several ribosomes translating the same RNA at the same time. In this pic, a gene is being transcribed. RNA Poly moving towards the right. The first part of the mRNA coming out is shown. Now we have a lot more of it that has emerged from RNA Poly. In this, as long as their is room for the ribosomes to assemble, they will. So a little bit of the 5' end of the nascent RNA is visible here, then we will immediately have a ribosome assemble and translate. We will have a steady stream of an mRNA being translated. |
|
|
Monosome |
1 ribosome associated on an mRNA molecule |
|
|
Polysome |
Multiple ribosomes associated on a single mRNA. Eukaryotes can have multiple ribosomes translating one mRNA, but almost always happens to prokaryotes. |
|
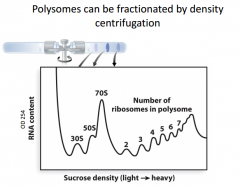
|
Experiment where we can measure the mRNA content (frequency of mRNA of a particular size). This is a density gradient experiment. We isolate ll the rna in a cell and centrifuge it in a sucrose gradient. The things at the top are going to be the lighter ones. The ones at the ends are going to be the heaviest ones. The highest frequencies are involved in the 70s unit (both ribosome units together). The next most common are 30s and 50s subunits. As we look at heavier components, each bump corresponds to an additional ribosome being on the mRNA. It is very frequent to have mRNA with many ribosomes, so they are more frequent. |
|
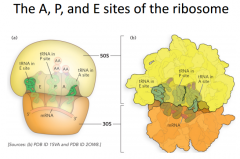
Most important sites of ribosome |
The 5' end of the mRNA is on the left. The 3' end is on the right. Each site is going to be most stable when it has a particular type of tRNA bound to it (thus they are tRNA binding sites). In the A site, we call it the aminoacyl tRNA site, meaning that a tRNA attached to 1 amino acid is the most favorable tRNA to be in that location. {That would be an example of the incoming tRNAs that are going to be going for the next codon in an mrna molecule. They come in bringing 1 aa, this aa will be transferred to the growing polypeptide chain and then finally once it has donated its aa, it will leave the ribosome} The second site is the P site (Peptydyl tRNA site). In most cases the most favorable steady interaction to have at this point is the tRNA with the growing polypeptide chain. E site is the exit site. This is where a tRNA is not attached to any aa. |
|
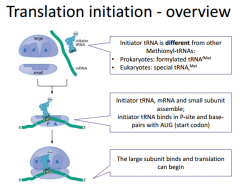
In prokaryotic translation, we will have an initiation phase, elongation and termination phase. Here: Initiation |
We will have our components (two ribosomal subunits, an mRNA molecule & an initiator tRNA.). The initiator tRNA is going to be different than the Met in the middle of the protein. It's going to be a Met in both Pro/Euk. We have to go through process where we get the mRNA loaded onto the ribosome so that it is actually in the correct reading frame so that the start codon on the mRNA is positioned to interact with the initiator tRNA. The initiator tRNA is going to bind within the P site. This is because the AUG start codon of the mRNA is directly beneath the P site. We have to set this up before we have the large subunit associate because we want to make sure we position the mRNA and tRNA in the right position because if we were to have the large subunit in complex at that point, we could potentially start translation at an inappropriate place. |
|
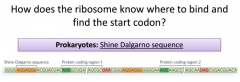
|
It's going to be a sequence that is slightly upstream from the AUG start. It works out so that once the SDS is recognized & bound by the 16s RNA and the small subunit, that will position the AUG start right underneath the P site. This shows how we can get 2 proteins from 1 mRNA. It's because they each have their own SDS. The 16s unit is going to be very critical for pairing with the SDS and making sure AUG start is positioned correctly. This base pairs within the SDS. The orientation of the 16s makes it so the AUG is right underneath P site. |
|
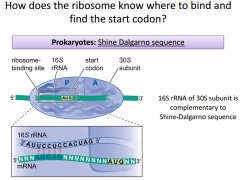
|
. |
|
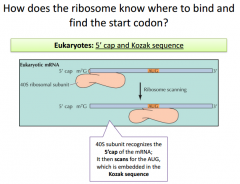
|
Similar mechanism with a consensus sequence we must pay attention to (Kozak Sequence). The KS is recognized by small subnuit in euk ribosomes. It binds at the 5' cap and scans along mrna looking for KS. KS is conserved within different organisms. KS incorporates the AUG start sequence somewhere. |