![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
44 Cards in this Set
- Front
- Back
|
Why is axon guidance so important |
Neural Circuitry is so precise and complex and it's almost all directed without trial and error because you don't want redundancy in CNS, so axon travel in CNS directed → stereotyped/error free |
|
|
Why does initial axon guidance happen when the brain is developing |
Distances are small, so pioneer neurons (Bale 1976) can easily navigate, then complexity is built up on top of that |
|
|
Is directed growth extrinsic or intrinsic? |
Rotate Mauthner Neurons in hindbrain 180deg → turns around when it reaches unrotated tissue → cues must be in the tissue! (Hibbard 1965). In fact global positional info across CNS. Although responsivity can vary = intrinsic reaction to the cues.... |
|
|
What is the Growth Cone? |
|
|
|
What are the two main structural parts of the growth cone |
Microfilaments - filopodia Microtubules - axon With intermediate zone → interaction between the two |
|
|
How to disrupt MTs/MFs and what happens |
Microfilaments (filopodia) Cytochalasin - depol Phalloidin - stabil
Microtubules (axon) Nocadazole - depol Taxol - stabil
depol → growth away; stabil → grow towards |
|
|
How influential are filopodia on neuron/growth cone |
Only one filopodia needed to touch laminin bead to redirect whole growth cone
|
|
|
What are the different shapes of the growth cone? |
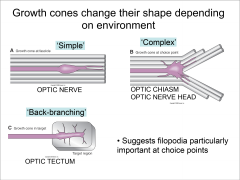
Simple - when growing normally Complex - Choice points Backbranching - When it reaches target |
|
|
What are the different types of substrate molecules (i.e. immobile, short-range cues) |
Need Permissive substrates - either:
These bind to SAMs (Integrins) or CAMs (hetero/homophilic) |
|
|
How do Pioneer neurons guide followers? |
Use selective adhesion with CAMs to bundle. Definitely needed ∵ if ablated → followers stall.
The specific CAMs for pioneer following i.e. bundling are:
Same neuron can express diff Fas at diff times - presumably to decide when to cross? Not sure how...
|
|
|
Are CAMs actually used for guidance? |
Not sure - definitely used for bundling (e.g. Fas in commissural or longitudinal growth) or growth (e.g. Tag1-NrCam responsible for midline crossing in retina).
Without Fas/Tag/NrCam → pathfinding becomes aberrant or can't grow past midline because substrate becomes repulsive, so used for guidance?
But deleting some single CAMs e.g. β integrins give few defects (Schwander et al 2004), so maybe not that critical. |
|
|
What are the classic Secreted, Long range guidance molecules? |
Netrin/DCC, Slit/Robo, Sema3a, Sema2 Responsivity to these can also vary at different times |
|
|
Netrin responsivity changes |
Netrin is attractive if Young, high cAMP Netrin is repulsive if Old, low cAMP (Shewan et al 2002) Also true with Slits, Semaphorins, Shh, BMP, Ephrins etc. |
|
|
3 diff examples of real life axon guidance from cocultures |
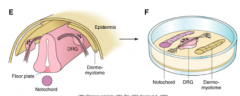
|
|
|
What attracts neurons to the midline? And which neurons do they attract/repel? |
Dorsal neural tube grows to floorplate in co-culture →Purify/fractionate → Netrin 1/2 (Serrafini et al 1994) → Dorsal neural tube grows to Netrin COS cells in co-culture (Kennedy et al 1994) If KO Netrin → commissural axons route wrong and don't function properly Ventral neurons e.g. Trochlear are repelled by Netrins ∵ diff combo receptors on growth cone e.g. Dcc-Unc5 = repulsive unlike Dcc-Dcc. |
|
|
Why would a neuron leave the midline and how do we know? |
Cut open the roof plate → see the whole neural plate flat (open book) → take out pre and post crossing and see attraction → post crossing lose attraction to floorplate AND repelled in coculture
Pre-crossing: Slit/Robo system silenced by Rig1/Robo3 (vert), or Comm (invert). Rig1/Robo3 directly silence, Comm downregulates Robo (so expression patterns mutually exclusive). Post-crossing: Slit/Robo silences DCC so no Netrin attraction.
The question still remains of whyRig1/Robo3 (vert), or Comm (invert) expression changes |
|
|
What happens in Robo / Comm mutants in inverts |
If Robo mutant → no suppression of attraction, so always crossing. Although why roundabout, not staying in the middle? If Comm mutant → never any suppression of netrin suppression, so always repulsed from midline |
|
|
What gradients used in AP direction for Axon guidance? |
Wnt = high Anteriorly Shh = high Posteriorly Does this contradict RA/FGF gradient in induction lectures? Not necessarily, because happens later.... |
|
|
Difference between Tadpole and Adult Xenopus at Optic Chiasm |
Tadpole - no ipsilateral neurons i.e. complete decussation Adult - some ipsilateral → allows binocularity because the receptive fields overlap |
|
|
How do you check if its the retina or the chiasm that changes from tadpole to adult? |
Take out ipsilateral adult neurons and heterochronically transplant them into a tadpole → grow contralaterally! So it must be the chiasm that is changing |
|
|
What changes from adult to tadpole chiasm? |
Ephrin B expression in chiasm changes. Expressed for 4 days → repels those neurons with EphB → ipsilaterally projectto allow crossing of certain neurons If Ephrin B knocked in → allows premature ipsilateralisation. EphB KO → severely reduced ipsilateral. Ephrin B receptor EphB is always present on the ipsilateral neurons (even in tadpole), and overlaps completely |
|
|
What are the main areas of activity with microfilaments in the filopodia? |
F actin assembly at tip (peripheral domain) F actin depolym at transition zone Also present in lamellipodia (webbing between filopodia fingers) |
|
|
What are the main properties of Microfilaments (size of subunit, growth properties, accessories that help growth) |
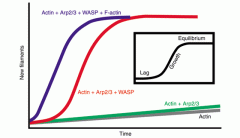
Actin = 48kDa subunit
Depends on concentration of actin units and presence of accessories |
|
|
How do cytochalasin and phalloidin work |
Cytochalasin - binds to ends of MFs and prevents polym. Can even sever filaments... Pholloidin - binds to MFs and lowers crit conc by preventing depol. |
|
|
What are some of the accessories of actin polym and how do they interact? What's the purpose of this? |
Cdc42 GTPase → binds N-Wasp → binds profilin and Arp2/3 → nucleates Actin This is one way in which actin polymerisation in filopodia is controlled |
|
|
Two models of leading edge migration
|
Cortical actin flow - actin growth pushes forward Membrane flow - vesicles add more membrane to one filopodia. Could be supported by actin flow because that's how vesicles travel... |
|
|
Clutch mechanism in the filopodia |
Jalin binds to integrins in filopodia which acts as a clutch because integrins bind to SAM/CAMs When unbound, the filopodia can slip backwards |
|
|
What is the structure of Microtubules and its properties |
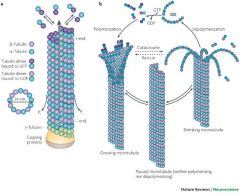
13 protofilaments vertically aligned in a tube All in the central domain - do seem to interact with the Actin |
|
|
Drugs pro/anti MT |
Nocadazol - inhibit pol by knocking out dynamic MTs by binding to them Taxol - binds directly to βtubulin and favours polymerisation |
|
|
How can you release drugs locally to affect cytoskeleton in growth cone |
Focal Laser Induced Photolysis on cage → releases taxol → turn in that direction because Taxol polymerises MTs Supports MT and MF interaction ∵ if MT stable → supports MF growth in that direction → filopodial growth |
|
|
What is Drebrin and what's its role? |
guidance cue → CDK → phosphorylate Drebrin → associate with Actin + binds EB3/tubulin complex → growth in that direction (Geraldo et al 2008) |
|
|
What does contact of filopodia with scleretome suggest? |
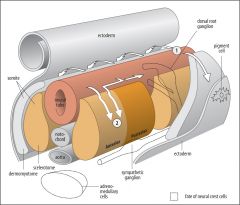
Anterior (Stimulatory) sceleretome contact → extend and then consolidate into neurite Posterior (Inhibitory) sceleretome contact → unable to extend due to structural instability Tip affects the shaft adhesion too (Steketee & Tosney 1999) |
|
|
What causes neurons to only grow through anterior chick somite segments? |
If you rotate somites so anterior now posterior, the neurons now grow through posterior, so its to do with intrinsic mols in somite. (Keynes, Stern 1984)
About 20 different molecules between A/P segments:
|
|
|
What do chick somites do to neurons? |
Posterior segments prevent axons coming through, only permitted through anterior segments |
|
|
What happens if you introduce proteins from posterior sceleretome to growth cones |
Use CHAPPS detergent → liposome → introduce proteins from P-sceleretome → collapses growth cone ∴ p-scleretome repulsive (Davies et al 1990) |
|
|
What is the Bonhoeffer stripe assay and what was it originally used to determine? |
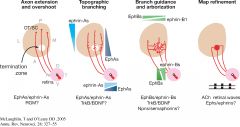
Strip assay show chick temporal RGC axons, choose to grow on lanes of anterior tectal membrane (not posterior), whereas nasal RGC grow on posterior. Not due to attractant on anterior tectal but instead to repellant by posterior - posterior membrane collapses. |
|
|
What are the gradients in the retina & colliculus/tectum |
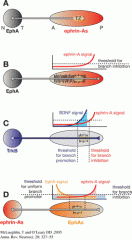
Varies from organism to organism, but generally Eph-Ephrin repulsioion that inhibits branching, but also others e.g. BDNF |
|
|
What are NoGo proteins? |
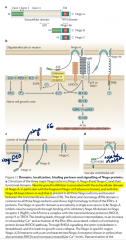
Nogo proteins = membrane proteins (basically a CAM) probably relevant for inhibiting growth/regeneration. 3 types - A,B,C Receptors on affected cell e.g. NGBR Also play a role in adult CNS stabilisation - possibly involving promotion of myelin growth. One Zebrafish example has it encouraging growth, weirdly. Not sure on intracellularmechanisms really: but probably uses Rho-GTPases / Ca2+, then can also beendocytosed -> cell body for decreased growth by lowering phosphorylation ofCREB and downregulation of growth genes. |
|
|
What 2 other receptors on the cell membrne interact with Netrin receptor DCC in modulating the ventral midline crossing mechanism? |
DSCAM - necessary and sufficient (Ly et al 2008) and APP (Rama et al 2012) |
|
|
What other chemoattractants might be involved with crossing at the ventral midline? |
Shh at midline = attractive BMP7/Draxins at Dorsal midline = repulsive. Important early on. |
|
|
How might the commissural decision be initially decided? |
"Those that are more sensitive to attractants cross the midline, whereas those axons that are more sensitive to the repellants remain ipsilateral" i.e. intrinsic attractiveness outweighing repulsivity to slit differentiates commissural vs non commissural (Dickson & Zou 2010) |
|
|
How might slit responsivity be turned back on at the midline? i.e. how might comm be turned off or rig1/robo3 be turned off |
Slit enrichment at the floorplate is so high that it finally turns on the repulsive response? i.e. there's a feedback from the repulsion mechanism to the repression of the repulsion? (Dickson & Zou 2010) Some have suggested intrinsic timing, but axons may spend diff times getting to midline so... |
|
|
What's the implication of midline changing responsivity for regular axon guidance? |
Normal axons that aren't crossing have guidepost cells which they have to be attracted to, then be repelled from. |
|
|
Are there are other repulsion systems at the midline apart from Slit-Robo? |
Shh and Sema3B/3F in mice (SHH upregulates cAMP which activates Semas). Also SCF in mice.
Shh in chicks too |