![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
107 Cards in this Set
- Front
- Back
|
The Cytoskeleton and its 3 main components |
Intricate network of protein filaments that extends throughout the cytoplasm Microtubules 25nm Microfilaments 7-9nm Intermediate filaments 10 nm |
|
|
Roles of the cytoskeleton |
Organelle/protein trafficking (vesicles are transported on the cytoskeleton) Cilia/flagella Mitosis/cytokinesis Muscle contraction Cell adhesion Cell migration Extravasation |
|
|
Structure of Microtubules |
Polymer of alpha and beta tubulin protein (basic subunit, polar - always ababab) Monomers are 55 kDa each - alpha tubulin binds permanently to GTP, beta tubulin can hydrolyze GTP After translation the alpha and beta tubulin quickly form a dimer (8nm) - very stable - beta subunit hydrolyzes GTP to grow protofilament (polymerization) One row of polymer is called a protofilament Protofilaments come together to form a hollow tube 25nm in diameter (MT) - we don't know how though Can be up to hundreds of micrometers long Can form flagella that stick out beyond the cell |
|
|
Arrangement of MT protofilaments |
Singlets - most common, not stable, 13 protofilaments forming single hollow tube of 25 nm Doublets - singlet of 13 + 10 protofilament B tubule, stable, found in cilia and flagella Triplet - singlet of 13 + 10 (B tubule) + 10 (C tubule), stable, found in basal bodies and centrioles |
|
|
Two types of microtubules |
Cytoplasmic (most) Axonemal (cilia and flagella) *has nothing to do with nerve axons, which have cytoplasmic microtubules |
|
|
Microtubule organization (MTOC) |
Polymerization around MTOC (microtubule organizing centre) Dynamic assembly Minus end of MT associated with MTOC (alpha tubulin) Plus end grows away from MTOC (beta tubulin) |
|
|
MTOCs and their microtubules |
Centrosome - cytoplasmic microtubules Spindle poles - mitotic spindle Basal body - axonemal microtubules |
|
|
Centrosome |
Major MTOC in animal cells (not found in plants) - contains centrioles: triplet MTs, don't directly polymerize singlet MTs, at 90 degrees to each other Singlets start in pericentriolar matrix (proteins around centrioles to help facilitate growth) - includes augmin complex, gamma tubulin
|
|
|
Centrioles |
Barrel shaped structures made up of triplet MTs Two barrels at 90 degrees to each other Around them are proteins that form and grow MT singlets (augmin, gamma tubulin, other pericentriolar material) Divide during mitosis to make mother and daughter centriole - daughter centriole is more naked, needs other proteins added to it |
|
|
Gamma tubulin ring complex (γTuRC) |
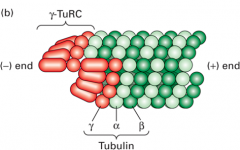
Complex of proteins in the pericentriolar space that provides nucleating sites for MTs, includes gamma tubulin monomers along with other proteins Binds to alpha end (minus end) which allows beta end (plus end) to grow |
|
|
Microtubule Formation (critical concentration) |
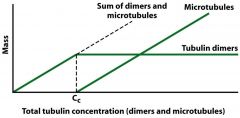
Increase the amount of alpha and beta tubulin At critical concentration (Cc) and a certain temperature the dimers will start polymerizing to MTs "Mass" = dimers + tubules (both increase) Amount of free tubulin will stay at critical concentration - extra dimers make the MT Humans don't shift temperature so it's not a factor for us, but lower temp = depolymerization, need higher Cc to polymerize Microtubules disassemble when chilled to 4 degrees C |
|
|
Polarity of Tubulin Polymerization |
Polymerization/depolymerization occurs faster at the + end Minus end has gamma tubulin ring complex, trapped Nucleating site accelerates initial polymerization - formed stable MTs can be nucleating centre (cilia and flagella doublets) - put nucleus in solution, add dimers, polymerizes over Cc |
|
|
Dynamics of Microtubules (dynamic instability graph) |
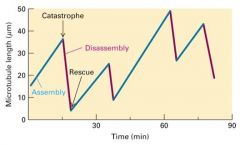
Oscillations in length is common - Dynamic instability Different areas of cell have different amounts of dimers - Microtubule grows, + end gets to area below Cc (point of catastrophe) - MT shrinks, + end gets to area above Cc (point of rescue) Assembly, Catastrophe, Disassembly, Rescue Not changing in size - MT is in equilibrium with Cc |
|
|
Polymerization and Depolymerization of microtubules (more specific) - lateral cohesion |
Both alpha and beta subunits must be in GTP form and above Cc - AB subunits added to + end (aka end with the beta subunits bound to GTP) FAST - as you add dimers, beta subunits hydrolyze GTP after polymerization (hydrolysis not needed for polymerization) - growing + end is neat: 13 protofilaments are tightly bound to each other (good lateral cohesion) - GDP bound beta subunits have less lateral cohesion between protofilaments - middle of the MT - when you run out of dimers at the + end, hydrolysis of GTP reaches the + end (GTP cap not maintained), which weakens lateral cohesion and smooth ends fray, beginning depolymerization |
|
|
Microtubule Disrupting Drugs |
- useful to study MTs Colchicine: depolymerizes MTs Taxol: stabilizes MTs - can be used as anti-cancer drug (mitosis requires depolymerization) |
|
|
Microtubule Associated Proteins (MAPs) |
- proteins bind to MT (middle) and alter stability and spacing of MTs - Stabilizing: MAPs coat the MT so it can't polymerize/depolymerize - MAPs have MT binding domain and projection domain (sticks out from MT, dictates how closely MTs can be packaged) - MAP2 and Tau are found in neurons, space MTs differently (Spacing affects structure and strength of nerve, transport) - MAPs are regulated: CDK phosphorylates them to promote disassembly (get rid of MTs to make room for mitotic spindle) |
|
|
+TIP MAPs |
Group of MAPs found on the + ends of MTs - caps the + end, stabilizes - can link the MT to something else EB1 is a +TIP - stabilizes + end - when EB1 present the MT is more likely to grow as well - moves along the crease inside the tubule (crevice where AB dimers loop around), possibly transporting things to the + end |
|
|
MT End Binding and Severing Proteins |
Proteins that sever MTs to speed up depolymerization - Kinesin 13 (ATPase) uses ATP hydrolysis to rip dimers off + end (able to depolymerize minus end except for gamma tubulin ring cap) - Stathmin binds tubulin dimers on the + end and promotes GTP hydrolysis in beta subunit, causes fraying of + end Regulation - Kinesin 13 regulated by amount of ATP available - Stathmin can be inactivated by phosphorylation |
|
|
How Kinesin was identified in Squid axons |
- inject radioactive amino acids into nerve ganglions, nerve makes radioactive proteins - radioactive proteins transported along nerve axon (MT) - isolate nerve and cut into discrete segments, isolate proteins from first segment (lyse cell and run on SDS PAGE) - Expose gel to X-ray film, black bands represent protein - inject and isolate at diff time points - shows that proteins move at set speeds and travel together - Cut protein bands from gel and sequence protein to identify it (kinesin!) |
|
|
Kinesin - basic structure |
+ end directed motor protein for MT - made of 2 heavy chains and two light chains - Heavy chains: head (ATPase, binds to MT), neck (linker), stalk (where light chains bind to tail) - Light chains: recognize cargo (vesicles, organelles), variable - most move to the + end of the MT |
|
|
Kinesin 13 |
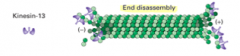
Involved in disassembling MT - uses ATP to rip apart dimers - no light chains - only has the heads of two heavy chains - not involved in transport |
|
|
Kinesin 5 |
Bipolar: 4 heavy chains, both ends have heads that can bind to MTs - involved in MT sliding |
|
|
Kinesin 1 |
Conventional kinesin (2 heavy and 2 variable light chains, heads move towards + end) - involved in organelle transport (anterograde movement, towards cell membrane) Head binds MT -> hydrolyzes ATP -> head and neck change conformation and walk along the MT's The two head domains start off 8 nm apart - when walking, one head goes past the other - heads move 16 nm for each ATP hydrolyzed |
|
|
Dynein |
Minus end directed motor protein for MT - 2 heavy chains: heads have ATPase activity, linker, stalk - Linker attaches head to stem, stem interacts with dynactin hetero complex (recognizes and binds to cargo) - ATP hydrolysis causes linker shape change that drives movement to minus end - dynactin also bound to MT to regulate movement (slow it down) |
|
|
Axoneme |
- structure of cilia and flagella - doublet MT outer ring, singlets in the middle - humans have 9+2 array (doublets + singlets) - outer doublets have A and B tubules (13 and 10 protofilaments), stable - made up of over 250 proteins, not just tubulin like MT - Nexin: links dimers together, ladder between doublet MT's - Axonemal dynein: stalk permanently bound to A tubule of doublets (not dynactin), dynein head reaching towards B tubule of next doublet |
|
|
Basal Body |
MTOC for cilia and flagella - 9 triplet MT's - directly responsible for polymerization of tubulin (unlike triplets in centriole) - Transition zone at cell surface, only A and B tubules pass through, making a doublet (cilia and flagella) - may have singlets, unrelate to singlets in axoneme - basal bodies often come in pairs, at 90 degrees to each other (cf. centrioles) |
|
|
Ciliary Beating |
Bending of cilia generated by MT sliding - Axonemal dynein (permanently bound to A tubule) head binds to B tubule of next doublet - motor walks towards minus end - MT's slide past each other, except they're bound to nexin so they bend instead! *bending is localized or else MT would rip itself apart |
|
|
Intraflagellar transport |
- cilia and flagella also used for cell signalling events, unrelated to bending - transport along the MT, involves kinesin (1?) and cytoplasmic dynein |
|
|
Non-motile primary cilium |
- almost all interphase cells have them - involved in signalling - Axonemal structure: doublet ring - receptors on the end of cilium, receives signals more efficiently - primary cilia often present in mammalian development (more important for embryos than adults) - mutate cilia (or transport along cilia) = developmental abnormalities |
|
|
Role of MT's in interphase of mitosis |
Centrosome duplication - centrioles replicate early by unknown mechanism - now 2 spindle poles (MTOC for mitotic spindle) instead of centrosome (diff MT network) - as centrioles replicate, get rid of interphase MT Interphase MT half life is 5 mins, vs. 15 secs in MT network in mitosis - MT have to capture all chromosomes and move to metaphase plate - Kinesin 13 and XMAP215 help with this |
|
|
XMAP215 |
Protein - stabilizing MAP (MT associated protein) - works with kinesin 13 (destabilizing protein and centrosomes to assemble MT's - suppresses catastrophes induced by kinesin 13 - inhibited by phosphorylation during mitosis, so that MT's are more dynamic, MTOCs can better nucleate MT's |
|
|
Mitotic Apparatus Spindle |
Composed of all MT - Kinetochore MT's: emit from poles, bind to chromosome at a kinetochore - Polar MT's: emit from poles but miss kinetochores, keep extending towards other pole - Astral MT's: emit from poles, polymerizing away from chromosomes to PM Mitotic Apparatus = Kinetochore + Polar + Astral Spindle = Kinetochore + Polar |
|
|
Centromere (and kinetochore) |
Attachment site for MT's - cohesin protein between chromosomes - kinetochore proteins mediate attachment of chromosomes to MT's Kinetochore: - two layers: inner and outer, different proteins - MT's captured by finger-like projections sticking out of kinetochore so that the + ends are still free - proteins bind to MT away from + end, not capped, free to polymerize and depolymerize |
|
|
Spindle Formation (MT dynamics of how chromosomes moved to metaphase plate) |
- MT's very unstable - chromosome captured from both sides Moving chromosome to the right: - depolymerize MT on the right side, polymerize MT on the left side - motor proteins: right side dynein pulls chromosome to the right (towards minus end), left side kinesin pushing chromosome to the right (towards free + end) *While polymerization and depolymerization themselves cause movement, motor proteins do most of the work |
|
|
Ndc80 |
One of the proteins used to capture chromosome at one kinetochore - binds to kinetochore and waits for tension from the MT on the other side - if no tension after a certain amount of time, Ndc80 is phosphorylated by a kinase resulting in weak MT interactions with kinetochore - ensures chromosomes are captured on both sides |
|
|
Anaphase A |
- requires MT shortening, especially the kinetochore MT's - brings chromosomes closer to the spindle pole - Depolymerization of + ends of kinetochore MT's - motor proteins help as well (dynein and kinesin?) (if you inhibit motor proteins depolymerization is enough to make Anaphase A happen) - Depolymerization of minus end shortens kinetochore MT, facilitated by motor proteins (kinesin and actin) |
|
|
Anaphase B |
- separation of the poles, requires motor proteins - chromosomal movements driven by motor proteins (not MT depolymerization like in Anaphase A) - Cell elongates during Anaphase A (Kinesin 5) - Polar MT's overlapping are antiparallel, with kinesin 5 between them (heads on both ends) - as kinesin 5 moves to + ends, the two MT's slide apart - when kinesin 5 gets to the end of MT's, polymerization often occurs so that kinesin can push the spindles further apart (cell makes sure no chromosomes in the middle still) - Poles pulled to the PM using Astral MT's: dynein is anchored to the PM and attached to the MT (minus end directed motor pulls spindle pole) - also depolymerization of + end of astral MT's so the MT's don't go through the PM |
|
|
Tracking chromosomal movements in Anaphase A |
- label spindle with fluorescent red - shine a strong light on fluorescent MT's at anaphase so that a section of spindle is bleached - position of bleached area stays the same but distance from bleached area to kinetochore shrinks -> + end depolymerizes (amount depolymerized at minus end is too small to be important) *Chromosomal movements are mostly driven by MT disassembly, even though motor proteins help out |
|
|
Function of Actin Filaments (Microfilaments) |
Cortical (associated with PM or underneath) - involved in cell shape ad movement - responsive to extracellular cues - reorganization of cytoskeleton as well |
|
|
Actin based structures |
Epithelial cells: microvilli, cell cortex, adherens belt Migrating cells: filopodia, lamellipodia, stress fibres Also phagocytosis, moving endocytic vesicles, contractile ring |
|
|
Structure of Actin (monomers and polymerized forms, polarity) |
Produced as a monomer g-actin (globular actin) - has ATP-binding cleft (and thus polarity) Polymerization is always in the same way because of polarity - F-actin is polymerized form (has plus and minus ends) - polymerize G-actin in ATP form and depolymerize in ADP form - you can depolymerize in ATP form through proteins (like kinesin 13 for MT) 1. Nucleation - need a nucleating centre before you start polymerizing 2. Elongation - polymerization at the + end is faster - add actin monomers until you hit Cc, then polymerization starts - G actin must be in ATP form 3. Steady State Assay to find minus end: If you add s1 myosin to an actin filament, it makes an arrowhead pattern that points to the minus end - also shows that actin has polarity - coating actin with s1 myosin stabilizes it (can be nucleating factor) |
|
|
Actin polymerization and Critical Concentration - Cc for minus and plus ends - what happens in relation to both Ccs |
Minus end critical concentration: 0.6 micromolar Plus end critical concentration: 0.12 micromolar (plus end polymerizes at lower concentrations) If concentration is: Above 0.6: polymerization at both ends 0.12 - 0.6: Treadmilling - polymerizing at plus end and depolymerizing on minus end (if you mark a monomer it'll look like it's moving to the minus end) Below 0.12: Depolymerization at both ends |
|
|
Regulation of Actin polymerization (proteins and actin-disrupting drugs) |
Thymosin: binds to and sequesters G-actin - lots of thymosin sitting in the cell - bound G-actin is no longer part of Cc reservoir - releases G-actin when Cc is needed Profilin: promotes polymerization by charging G-ADP into G-ATP actin - cell turns on profilin when it wants polymerization Cofilin: enhances depolymerization - breaks monomers off at the minus end CapZ: binds to the + end and stabilizes it Tropomodulin: binds to the minus end and stabilizes it Drugs! Cytochalasin: depolymerizes actin filaments Phalloidin: stabilizes actin filaments - both can help with actin research, add fluorescence to it |
|
|
Assembly and Branching of Filaments - proteins involved - Listeria example - endocytosis example - phagocytosis example |
Formin: assembles unbranched filaments at the + end - dimer, sits on the + end - regulated by Rho GTPase that also must be activated (Formin is a cap if inactive) Arp2/3: makes a branch point (like a new nucleating centre) - regulated by CDC42 (GTPase) working with WASp - CDC42 must be in GTP form to be active and activate Arp2/3 Listeria - parasite with proteins on its cell surface that activate Arp2/3, gets inside your cells - fast actin polymerization at parasite's tail end, moving around the cell - parasite bumps into PM and tries to break through it by force to get to neighbouring cells Receptor-mediated endocytosis - actin is polymerized (dependent on Arp2/3) and pulls the membrane inwards to start formation of vesicle (and then dynamin breaks it off) Phagocytosis (ex. leukocytes) - actin polymerizes and pushes PM out and around pathogen to encircle it |
|
|
Actin-binding proteins - support cellular structures |
Fimbrin and Alpha Actinin: bundle actin proteins together into stiff, stable parallel structure - makes unbranched microfilaments - these bundles in microvilli (diff from cilia, they can't move) Spectrin and Filamin: cause more links in branched networks, adding to complexity and helping form crosslinks for tighter networks Ankyrin: anchors actin network to PM, often with PM proteins like Band3 Ezrin: links actin to PM protein in microvilli - actin pushes against PM outward Dystrophin: important in muscle cells. Actin contracts, pulls on dystrophin, which pulls on glycoprotein complex -> pulls tendon -> pulls bone - muscular dystrophy: mutations in dystrophin so it doesn't bind actin properly - cell contracts but doesn't pull properly on ECM so it works harder and damages itself - eventually degeneration, cells don't grow back |
|
|
Myosin |
- similar to kinesin and dynein - has heavy chains (ATPase head that binds to actin and moves to + end, neck, tail that binds cargo) and light chains (bind at the neck and regulate speed of movement) - need two heads to move - + end directed |
|
|
Types of Myosin (3) |
Type I Myosin (Membrane association) - does not form a dimer - binds to membranes (microvilli) - mostly holds things in place - can be involved in endocytosis - can move if there's enough ATP available Type II Myosin (Contraction) - forms thick filament involved in contraction - usually found as a dimer (two heads) Type V Myosin (Transport) - variable tails - involved in most organelle/vesicle transport |
|
|
Sliding filament assay |
- used to detect myosin-powered movement Process: - take myosin heads and add to glass slide - add labeled actin, which will bind to myosin in the presence of ATP - you'll get movement, you can measure step size, speed, effect of diff light chains and myosin types, function of ATP Learned: - myosin V heads step head over head (like kinesin), each head steps 72 nm - longer myosin II neck = more light chains can bind = faster movement |
|
|
Conformational Changes of Myosin (Actin-Myosin ATP cycle) Rigor mortis |
- Rigor state: myosin bound to actin, no ATP - when ATP present, binds to myosin head, myosin head releases actin filament - ATP hydrolyzed, ADP and phosphate not released from myosin head - ^causes myosin head to bend and bind to actin monomer towards its + end - Powerstroke: phosphate released, head and neck change and actin molecule is moved - ADP released, back in rigor state Rigor mortis - stiff muscles, no ATP that triggers myosin to let go of actin, stuck in rigor state - myosin sticks to actin until muscles start to decay |
|
|
Skeletal Muscle Sarcomere |
Z discs are vertical pillars - actin microfilaments extend from Z discs - actin is aligned so the + ends are at the Z discs Myosin filaments have their head domains pointed at Z discs (+ end) Powerstroke pulls Z discs closer together - the sarcomere shrinks - A band (myosin) does not change size |
|
|
Stabilizing and scaffolding proteins that bind to skeletal muscle (sarcomere) |
Actin stuff CapZ: caps + end of actin Tropomodulin: caps minus end of actin Nebulin: wraps actin to stabilize, but still allows myosin to bind Myosin stuff Titin: holds myosin II filaments in the middle of the sarcomere - a little elastic - as sarcomere contracts, titin is compressed - as sarcomere relaxes, titin pushes sarcomeres apart a bit with elasticity |
|
|
Ca2+ release from sarcoplasmic reticulum |
T tubules are extensions of PM that contact the SR Nerve impulse -> depolarization -> electrical change comes down nerve, T tubule, triggers Ca2+ release from SR from voltage-gated channels -> contraction occurs quickly -> Ca2+ immediately pumped back into SR with ATP pump Whole cycle is about 30 msec |
|
|
How Ca2+ causes muscle contraction |
Troponin and tropomyosin are bound to actin filament - without Ca2+, tropomyosin coats the actin in a way that covers the myosin binding site Troponin binds Ca2+ -> Troponin changes shape and shifts Tropomyosin position -> Myosin binding site exposed on actin -> myosin binds to actin and contraction can occur |
|
|
Actin and Myosin in Non-Skeletal Muscle Cells (list 5) |
1. Contractile ring in cytokinesis 2. Myosin phosphorylation in smooth muscle contraction 3. Vesicle transport (Myosin V) 4. Cytoplasmic streaming 5. Cell Migration - stress fibres |
|
|
Localization of Myosin I and II during cytokinesis (contractile ring) |
Contractile ring is a looped sarcomere-like structure - has actin and myosin Myosin II: thick filament, found in cleavage furrow Myosin I: involved in holding PM at the poles so cell doesn't collapse Ring shrinks (and eventually disappears?) via simultaneous contraction and depolymerization of the sarcomere |
|
|
Regulation of Smooth muscle contraction via Myosin phosphorylation |
Smooth muscle works slower and the contractions last longer (small intestine) - myosin not always functional and active, need to phosphorylate it to turn it on (also need ATP and other stuff) |
|
|
Role of actin and myosin V in vesicle transport (ex. yeast bud) |
You need to get everything needed for a new cell into the bud! Vesicles, mRNA, etc. - actin network forms where + end leads into bud - because myosin is + end directed motor, it enters the bud - use formin to help polymerization of actin at its + end - actin and tubulin (MTs) work together to move the nucleus into the bud |
|
|
Cytoplasmic streaming |
Plant cells depend on diffusion, and diffusion works better with cytoplasmic streaming - cortical actin in plant cells forms train track around the cell - myosin molecules move around the actin network and cause the whole cytoplasm to constantly move around |
|
|
Stress fibres
|
long actin fibre bundles interacting with myosin and other regulatory molecules - they are contractile |
|
|
Chemotaxis |
Signalling molecule attractant - needs appropriate receptor for the signal, ideally receptors all over cell surface so it can move any way it wants - cell gets chemotactic agent and alters its cytoskeleton via leading edge signalling cascade - once cell migration begins, start polymerizing actin and myosin to form the leading edge |
|
|
4 steps of cell movement |
Transmembrane proteins link cell to ECM - focal adhesions at either ends of stress fibres - cell sits on a substrate with focal adhesions, receives chemotactic signal 1. Extension - actin polymerization at leading edge triggered - actin pushes out PM at leading edge forming lamellipodium 2. Adhesion - if cell finds a favourable substrate to stick to in front of it, it forms a new focal adhesion and thus a new stress fibre 3. Translocation - actin and myosin contract, moves the cell body forward 4. De-adhesion and endocytic recycling - de-adhesion at the back end, get rid of posterior stress fibre |
|
|
Role of CDC42, Rac, Rho, and actin in cell migration - family of proteins - non-motile functions of proteins - test with dominant active forms |
CDC42, Rac and Rho are "Rho proteins" - members of Ras superfamily of small GTPases - GTP active forms can trigger effector proteins Actin polymerization (non-motile cell) requires Rho GTPases - Rho activates Formin - CDC42 activates Arp2/3 Dominant active CDC42: protrusions coming out of cell, formation of actin bundles pushing the PM forward to form philopodia (small lamellipodia) Dominant active Rac: networking and branching of actin, push out large chunks of membrane to form lots of lamellipodia Dominant active Rho: lots of focal adhesions, actin and myosin organized into stress fibres |
|
|
Coordinate regulation of CDC42, Rac and Rho in cell migration |
- chemoattractive molecule binds to receptor, activates GTPases - first CDC42 triggers philopodia formation, they're sent out towards the signal - growth factor activates Rac at the same time - actin networks at the same time as actin bundles, lamellipodia sent out to look for a place to put a new focal adhesion - when philopodia find a place to stick, Rho is activated - interacts with myosin and forms stress fibres at the back of the cell, contraction and movement |
|
|
Scratch Assay |
If you grow cells to confluence and scratch a clear area, cells will move to close the wound fairly quickly If you take cells with dominant negative forms of CDC42, Rac and Rho - migration doesn't work, wound closure inhibited - tells us that the proteins need to work together and trigger each other |
|
|
Main function of intermediate filaments Main types of intermediate filaments |
Main function is to hold things together, tensile strength - dynamic Types Keratin: epithelial cells (living and dead), holds skin together Desmin: muscle, supports sarcomeres (along with dystrophin) - holds muscles together (they exert force on each other) Vimentin: mesenchymal cells (migrating) Neurofilaments: supports axons Lamin: not cell specific, supports nuclear envelope (keeps it open and allows transcription and stuff to occur) |
|
|
Structure of intermediate filaments
|
Basic building block is a tetramer - two elongated dimers of protein come together (not globular like MT and MF) - symmetrical, no polarity (so no polarity after polymerization) |
|
|
Lamin A |
- found in nucleus, support nuclear membrane - CDK phosphorylates a serine residue on lamin - induces disassembly and prevents reassembly of IFs - breaks apart nucleus envelope - essential for mitosis (if you mutate the serine residue, mitosis won't occur) |
|
|
Intermediate filament associated proteins |
- only organizational proteins have been identified (no motors, caps, severing) Plectin: links IFs to MTs - IFAP and MAP - strengthens network Lamin Associated Polypeptide (LAP2) - Links Lamin A/B network to nuclear lamina |
|
|
Functions of IF in membranes |
Provides structural support necessary for cell shape, keeps nucleus open - can bind to membranes (like actin) Lamins support nuclear membrane - linked to nuclear lamin via LAP2 Vimentin links to ankyrin to support PM (ankyrin can also link to actin) |
|
|
Keratin Mutations |
Transgenic mice with non functional keratin: skin cells can't hold together properly - can rip epidermis off the dermis Normally the IFs are anchored to desmosomes and hemidesmosomes (molecules involved in cell adhesion) |
|
|
IF junctions: desmosomes and hemidesmosomes (keratin) |
Keratin links from one desmosome to another, a rope from inside one cell to the inside of another Desmosomes link cells to other cells Hemidesmosomes link cells to ECM |
|
|
Cell Junctions and Cell Adhesions (4 types) - and polarity in epithelial cells |
1. Tight Junctions 2. Gap junctions 3. Cell-cell adhesions 4. Cell-ECM adhesions Epithelial cells: junctions have polarity - apical surface is to the outside |
|
|
Gap Junctions |
Channels that exist between neighbouring cells Connexin protein: transmembrane, 6 of them form a connexon (hole in PM) Connexons on two cells join and form a gap junction - ions can get through easily (electrical charge is important in epithelial cells) Danger in linking cells - if you damage the sheet, close gap junctions quick before cytoplasm leaks into other cells - Calcium (secondary messenger) enters from outside the cell, binds to gap junctions and closes them |
|
|
Tight Junctions |
Glue PMs together via a ring by the tops of epithelial cells - usually formed towards apical surface - prevents paracellular transport - also involved in signalling - involves a row of Occludin, Claudin, and JAM proteins that bind to similar proteins on the other cell and glue the membranes together - bound to actin |
|
|
Cell adhesion molecules (CAMs) |
Transmembrane molecules for cell-cell adhesion or cell-ECM adhesion - linked to cytoskeleton, provide strength Either involved in Homophilic or Heterophilic interactions - Homophilic: Need identical partner on other cell to bind (cadherin, IG superfamily) - cell-cell only - Heterophilic: bind to a diff molecule (integrin, selectin) - cell-cell or cell-ECM |
|
|
Cadherin |
Over 40 types Homophilic adhesion - needs to bind to the same type of cadherin, must be cell-cell Family members: desmoglein and desmocollin, E-cadherin Single transmembrane domain, cytosolic C-terminal tail associated with cytoskeleton (thru adapter proteins) - bound to IFs - they're desmosomes - can bind to actin (possibility for contraction) - adherens junction - adherens junction can form belt around apical surface of cell: circumferential belts (actin) - if cells ripped apart, adapter proteins can signal nucleus saying that adherens junction is broken Ca2+ dependent - ECM domain (N-terminus?) needs Ca2+ to bind (it's usually around anyways) |
|
|
N-CAMs (IG Superfamily) |
Homophilic interactions - unlike cadherins, don't need EC calcium present for them to bind (experiments with and without calcium can determine whether cadherins or Ig superfamily are present) - no cytoplasmic domain, not bound to cytoskeleton, can't signal the cell - some may not have transmembrane domain, just GPI anchor |
|
|
Selectins |
P-selectin recognizes certain oligosaccharide - helps identify cells Heterophilic interactions |
|
|
Extravasation |
involves heterophilic interactions, cell-cell adhesion Leukocytes leave the bloodstream to fight an infection - endothelial cells line bloodstream, tight junctions keep blood in - Infection outside of blood signals to endothelial cells - Endothelial cells secrete P-selectins on PM, specific for sugars on the leukocyte - they grab the leukocyte and slow it down - Integrins on leukocyte become active and recognize ICAMs (RGD recognition) on endothelial cells to tightly bind leukocyte to PM *leukocyte must be bound to P-selectin for integrin to bind to ICAM |
|
|
Integrin |
Heterodimer (alpha and beta subunit) Heterophilic interactions (any kind) - bind to amino acids RGD (arginine, lysine, aspartic acid) in core of integrin - recognizes amino acids around RGD, giving integrins their substrate specificity - integrin must be activated internally by the cell - linked to cytoskeleton so that when it binds to substrate, cell knows and turns on signalling pathways a1b1 integrin binds collagen a5b1 integrin binds fibronectin a6b1 integrin binds laminin - membrane projection, forms focal adhesion Migratory cells: - bind to actin to form stress fibre (linked with focal adhesion formation above) Non-migratory cells: - bind to IF to for hemidesmosome (other side to laminin) |
|
|
Extracellular Matrix - function - major components |
Function: - allows binding of cells in tissues - allows/inhibits cell movement - modulates ligand receptor interaction Major components: - Collagen: large, structural - Fibronectin and Laminin: multi-adhesive, allows migration - Proteoglycans: have large sugar residues, can absorb a lot of water -> tissue resiliency (ex. eye) |
|
|
Cytoskeleton and Adhesion in Epithelial cells |
Microtubules - coming from MTOC Microfilaments (actin) - cortical, supports PM - actin bundles form microvilli - possible adherens junctions between cells with cadherins (signalling) - tight junctions Intermediate filaments - nucleus supported by Lamin - desmosomes (cadherins) between cells - hemidesmosomes (integrins) between cell and ECM *Non-migratory |
|
|
Cytoskeleton and Adhesion in Migratory cells |
- unlikely to have cell-cell adhesion (no desmosomes, adherens junctions, tight/gap junctions) - would have cell-ECM stuff tho (hemidesmosomes?) Microtubules - present, from MTOC Microfilaments (actin) - cortical, supporting PM - philopodia, lamellipodia, stress fibres, focal adhesions Intermediate filaments - Lamin supporting nuclear envelope |
|
|
Basic components of biomembranes |
Lipids, sterols and proteins |
|
|
Relationship between melting point, chain length of fatty acids and their unsaturation |
Chain length increase = melting point increase Unsaturation increase = melting point decrease |
|
|
Fatty acid |
Long hydrocarbon chain attached to polar carboxyl head group - amphipathic - Cx:y where x = number of C molecules, y = number of double bonds |
|
|
Properties of Biomembranes (4) |
1. Fluid - rapid lateral diffusion - dependent on fatty acids (lengths, saturation), presence of sterols (unpredictable), heat - transmembrane proteins connected to cytoskeleton fix parts of the membrane 2. Closed compartments - in PM, cytosolic face is internal - in Vesicles, cytosolic face is external 3. Semi-permeable - cannot pass if large, charged, hydrophilic 4. Asymmetric - slow transversion between leaflets - phospholipid composition different - proteins embedded in fixed orientation - carbohydrates only found on exoplasmic face |
|
|
FRAP |
Fluorescent Recovery After Photobleaching
- label proteins on outer leaflet of PM - bleach area with a laser (Time 0) - look for fluorescence recovery - measures rate of lateral diffusion - affected by heat - diffusion 10x slower in PMs than pure bilayers, up to 50% immobile |
|
|
Membrane Proteins (3 types) |
*all are asymmetric 1. Integral 2. Lipid-linked 3. Peripheral |
|
|
Integral Membrane proteins |
3 distinct domains: cytoplasmic, transmembrane, exoplasmic - transmembrane domain has hydrophobic secondary or tertiary structures: a helice or b barrel - mostly glycosylated in exoplasmic domains (glycosylation happens in ER and Golgi) |
|
|
Alpha helix
|
- approx. 20-25 amino acids - hydrophobic - often charged amino acids (lys and Arg) near cytosolic side, they interact with polar heads of phospholipids to further anchor protein to the membrane (also means it is less likely to flip leaflets) |
|
|
Lipid-linked proteins |
- no transmembrane domain, directly linked to lipid bilayer - good lateral movement 3 Important Linkages: 1. Acylation - Attaches N-terminal glycine to cytoplasmic surface 2. Prenylation - attaches C-terminal cysteine to cytoplasmic surface (ex. Ras) 3. GPI Anchor - links protein to specific residues on exoplasmic surface - ex. N-CAM - requires phosphoethanolamine and phosphatidylinositol |
|
|
Peripheral Proteins |
- bound to another membrane protein through non-covalent interactions (ionic, H-bond, protein-protein, van der Waals)
- can be bound on either side depending on what it is, asymmetric - ex. Adapter proteins with cadherin, Dystrophin, Ankyrin - can help link bilayer and ECM to cytoskeleton |
|
|
Topogenic sequences |
Sequences identified by other proteins after translation, tell new protein where to go to help with more folding 1. N-terminal (cleaved) signal sequence 2. Stop-transfer/membrane anchor sequence (STA) -> back to cytosol 3. Signal-anchor - internal (uncleaved) sequence (SA) 4. Hydrophobic C-terminus Types of Proteins made by topogenic sequences: - Type I: N in ER lumen, C in cytosol, STA transmembrane - Type II: N in cytosol, C in ER lumen, SA transmembrane - Type III: N in ER lumen, C in cytosol, SA transmembrane - Type IV - GPI linked protein - Tail-anchored protein: N in cytosol, C in ER membrane |
|
|
Tail anchored proteins |
Topogenic sequences: hydrophobic C-terminal domain Get3 protein recognizes hydrophobic tail and brings protein to ER membrane - Get1 and Get2 (in ER membrane, ATP hydrolysis) insert hydrophobic tail into the ER membrane - rest of protein stays in the cytosol - tail now anchored in ER membrane |
|
|
Type I proteins synthesis |

Topogenic sequences: N terminal signal sequence, STA - signal recognition particle recognizes N-terminal signal sequence, takes protein to ER (translocon) while translation still going - translation now happening in ER lumen, N-terminal signal sequence cleaved - STA sequence recognized: protein translated into cytosol, STA itself becomes transmembrane domain - end up with N-terminal domain in the ER lumen and C-terminal domain in the cytosol |
|
|
Type II and III proteins synthesis |
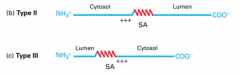
Topogenic sequences: SA - also depends on location of positively charged amino acids - SA sequence brings protein to ER (SA becomes transmembrane domain) - If charged amino acids on the N-terminal side around SA, N stays in the cytosol and C ends up in lumen - Type II - If no charged amino acids on N side then there will be on C side, N put into lumen and C ends up in cytosol - Type III |
|
|
Type IV protein synthesis |
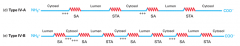
Topogenic seqeunces: SA and STA alternating Type IVA: alternate SA and STA, start with SA and charged AA on N side - N in cytosol Type IVB: SA with charged AA on C side - N in lumen - need another SA with N side AA to get next section into lumen |
|
|
Passive Diffusion - partition coefficient |
- small and uncharged molecules get through Partition coefficient: measure of preference of molecule to partition into a hydrophobic environment Higher K = more lipid soluble = can undergo passive diffusion more easily |
|
|
Facilitated Transport |
- driven by concentration gradient - Types: pores channels, gates, uniporters - hydrophilic stuff moves through protein lined pathway through PM - faster than passive diffusion - specific - saturable ex. GLUT1 (glucose uniporter) |
|
|
K+ Resting Channel (size selectivity) (a pore) |
- K+ must release water molecules (hydration shell) to enter the channel - oxygen molecules in the pore with the same spacing - result: not much energy difference in going through the channel for K+, so its driven by its concentration gradient - Diff ions have diff sized hydration shells = makes the pores specific - can lead to electric potential across the membrane without any further input of energy |
|
|
Facilitated Transport: Gates |
- normally closed, open when needed Ligand-gated: only opens when ligand binds to it (GLUT1) Voltage-gated: opens when membrane potential changes (Ca2+ release from SR for muscle contraction) |
|
|
Active Transport (Primary and Secondary)
|
Primary active transport: ATP-powered "pumps" - P-class - H+, Na+, K+, Ca2+ - V-class - H+ only (endosome acidification) - F-class - H+ only - ABC Type - many small molecules Secondary active transport - need primary active transport first - energy from that system can be coupled to moving other stuff against their gradients (ex. symporters and antiporters - one molecule with gradient and one against) |
|
|
P-class Pump, Muscle Ca2+ ATPase |
- pumps calcium back into SR as soon as it is released (by voltage-gated channel) 2 calciums out of cytosol for every ATP hydrolysis with the pump |
|
|
P-class Pump, Na+/K+ ATPase |
3Na out of the cytosol and 2K into the cytosol for every ATP hydrolysis - creates salty banana: high sodium outside and high potassium inside the cell |
|
|
Na+ Glucose Symporter |
Most cells have K+ resting channel and Na/K ATPase -> high sodium and positive charge outside cell - 2Na+ down gradient, 1 glucose against gadient |
|
|
Co-transport in epithelial cells |
- cells have low Na and high K (Na/K pump) - inside of cell is negative Apical membrane - 2 Na/Glu symporter (secondary active transport) Basolateral membrane - Na/K pump sets up gradient (primary active transport) - GLUT2 (uniporter, facilitated diffusion) |