![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
50 Cards in this Set
- Front
- Back
|
4 .17 De rol van de mediale temporale regio
Van alle hersenstructuren die geassocieerd worden met de geheugenwerking zijn de temporale lobben het meest genoemd.
|
post mortem neuropathologische bevindingen van een bilaterale mediaal temporale verwerking waar bij ook de formatio hippocampi getroffen bleek.
|
|
|
De klassieke gevalsstudie H.M. illustreerde de samenhang tussen |
hippocampus en het geheugenfungeren. |
|
|
De oorzaak van H.M.'s fenomenale geheugendeficit werd toegeschreven aan |
de bilaterale verwijdering van de hippocampus.
|
|
|
De hippocampus is een ongeveer acht centimeter lange structuur van primitieve cortex gelegen aan |
de mediale zijde van de temporale lob.
Het woord 'hippocampus' betekent 'zeepaardje' en werd wegens de fysieke gelijkende naam voor deze anatomische structuur. |
|
|
Het geval H.M. leert dat de hippocampus een belangrijke rol speelt bij de vorming van het ………..geheugen. |
declaratief geheugen.
H.M . bleek immers niet meer in staat episodische en semantische informatie op te slaan.
Anderzijds bleek zijn geheugen voor procedurele informatie ongeschonden zodat we moeten besluiten dat een intacte hippocampu niet noodzakelijk is voor het impliciete non-declaratieve geheugen.
|
|
|
Het geval H.M. leert ons bovendien dat de hippocampus niet de plaats is waar |
onze geheugeninhouden opgeslagen liggen.
Indien dit wel het geval was, zou H.M. ook alle voordien aangeleerde kennis verloren zijn, wat duidelijk niet het geval was.
|
|
|
De hippocampale regio dient veeleer als een |
tijdelijke bewaarplaats voor nieuw aangeleerde informatie die later in andere corticale regio's zal worden bewaard.
Ook andere gevallen met een bilaterale beschadiging van de hippocampus vertonen een amnestisch syndroom, zij het niet zo uitgesproken als H.M. (Squire, 1992b ; Zola-Mor gan, Squire & Amaral, 1986).
|
|
|
We mogen evenwel niet vergeten dat H.M.'s amnesie niet enkel werd veroorzaakt door hippocampale schade, maar dat bij deze patiënt ook andere met de hippocampus geassocieerde structuren zoals ….. werden verwijderd. |
de entorhinale, perirhinale en para hippocampale cortices werden verwijderd (Squire, 1992b).
|
|
|
Patiënten met unilaterale laesies van de hippocampus hebben problemen met het opslaan van nieu we informatie, maar vertonen niet de ernstige amnesie van mensen als
Unilaterale laesies bieden aanwijzingen voor |
een modaliteitsspecifieke taakverdeling waarbij de linkerhippocarnpus vooral betrokken is bij de opslag van verbaal materiaal en de rechterhippocampus instaat voor de opslag van visueel en spatieel materiaal
(Kolb & Whishaw, 1990; Tranel, 1991). |
|
|
Het hippocampaal systeem (formatie hippocampi) bestaat uit de
C S E G |
-cornu ammonis (de eigenlijke hippocampus ), -subiculum en de -entorhinale cortex, en de -gyrus dentatus .
|
|
|
-De cornu anunonis bestaat uit -Het subiculum vormt de -De entorhinale cortex is het meest -De gyrus dentatus bedekt de |
-De cornu anunonis bestaat uit verschillende velden (CA I , CA2 en CA3) met een drielagige cortex waaronder piramidale cellen.
-Het subiculum vormt de overgang tussen de zeslagige cortex van de entorhinale area en de primitievere drielagige hippocampale cortex.
-De entorhinale cortex is het meest mediale deel van de gyrus parahippocampus en de belangrijkste weg waarlang prikkels de hippocampus bereiken .
De entorhinale cortex is het meest informatie van vele hersenstruc turen zoals directe projectie van de orbitofrontale en olfactorische cortex, de amygdala, septale kernen , thalarmus en dopaminerge, erotonerge en noradrenerge celgroepen van de middenhersenen. Maar ook van de insulaire en cingulate cortex en de gyrus temporali superior.
De meest belangrijke input naar de entorhinale cortex komt op indirecte wijze vanuit alle corticale gebieden door middel van een multisynaptisch transcorticale route doorheen de a sociatieareas naar de temporale lob via de perirhinale en parahippocampale cortex. Virtueel alle perceptuele , gedrag matige en cognitieve bewerkingen van de cerebrale cortex worden op die manier via de entorhinale cortex naar het hippocampale verwerkingscircuit gestuurd .
Beschadiging van de entorhinale cortex verstoort dan ook de hippocampale functie (degeneratie van de entorhi nale area is één van de eerste pathologische tekenen van de ziekte van Alzheimer waarvan een ernstig geheugendeficit het meest op de voorgrond tredende symptoom is).
-De gyrus dentatus bedekt de hippo campus als de kroon van een tand , waaraa n deze anatomische constructie haar naam ontleende. In de gyrus dentatus wordt de piramidale cellaag van de drielagige cortex vervangen door een granulaire cellaag.
|
|
|
Het neuronaal circuit van het hippocampaal complex blijkt goed georganiseerd (Kennedy & Marder, 1992).
Figuur 4.11. toont een schema van deze organisatie.
|
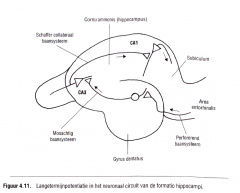
|
|
|
De entorhinale cortex ontvangt input van vele corticale associatiearea en biedt de formatio hippo campi een belangrijke bron van gegevens waaruit een herinnering kan worden opgebouwd.
De structuren van het hippocampaal complex hebben immers toegang tot en invloed over signalen van uit |
het gehele brein .
Het systeem bevindt zich in een sleutelpositie om een multimodale voorstelling op te bouwen die visuele, auditieve en somatosen sorische informatie integreert.
|
|
|
De input naar de hippocampus wordt verzorgd door axonen die vertrekken vanuit de piramidale neuronen in de entorhinale cortex.
Deze piramidale axonen kruisen de hippocampale fissuur als een diffuse zenuwbundel en projecteren naar de granulaire cellen van de gyrusdentatus.
Men noemt deze projectie |
het perforerend baansysteem ('perforant pathway ').
|
|
|
De axonen van de granulaire cellen van de gyrus dentatus, ook wel mosachtige ('mossy') vezels genoemd , leiden de prikkels naar de dendrieten van de piramidale cellen in het cornu ammonisveld CA3.
De piramidale axonen van CA3 projecteren op hun beurt naar |
de piramidale dendrieten van het cornu ammonisveld CA l via het collateraal baansyteem van Schaffer. |
|
|
De axonen van de CAI-piramidale cellen projecteren naar het subiculum en van daaruit terug naar de |
entorhinale cortex.
Op die manier ontstaat een circuit van prikkelgeleiding binnen het hippocampaal systeem.
|
|
|
Naast het bestaan van dit hippocampaal circuit is het belangrijk op te merken dat alle neuronen van de formatio hippocampi (de neuronen van de gyrusdentatus uitgezonderd) tevens axonen naar andere regio' van de cerebrale cortex versturen.
Het hippocampaJe systeem is derhalve |
ideaal geplaatst om de geheugenfunctie in de ver preide hersenregio's te controleren .
|
|
|
De axonen die de hippocampus verlaten , vormen samen de toroix en ontspringen vooral van uit de piramidale neuronen van de area CA 1 en het subiculum (Figuur 4.12.).
De fornicale vezels vanuit CA 1 eindigen vooral op neuronen van de septale kernen (basale voorher enen) die ook projecties terugzenden. |
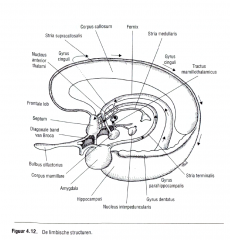
|
|
|
Die van het subiculum projecteren vooral naar de corpora mamillaria van de hypothalamus.
Via axonen die samen het mamillothalaroisch baansysteem vormen, zenden de corpora mamillaria de signalen die hen via de fornix bereiken, naar |
de anterieure kernen van de thalamus.
|
|
|
De anterieure thalamuskernen voorzien in een thalamische input naar de cortex via de gyrus cinguli aan de mediale oppervlakte van |
de hemisfeer.
|
|
|
Vanuit de gyrus cinguli vertrekken signalen via de cinguli naar de gyrus parahippocampus en komen |
opnieuw in de formatio hippocampali terecht.
Deze anatomische lus werd voor het eerst door James Papez beschreven. |
|
|
Vooral de ontdekking dat hippocampale neuronen de sterkte van hun reactie kunnen aanpassen in functie van voorgaande informatie, bleek een belangrijke verwijzing naar
|
de fysiologische basis van expliciet geheugenfunctioneren.
Dit fenomeen wordt langetermijnpotentiatie genoemd en verwijst naar een toegenomen neuronale reactiviteit op een gewone (standaard) stimulatie in een pecifieke hersenregio naar aanleiding van een korte, intense elektrische stimulatie van enkele afferente neuronen die naar deze regio projecteren (Bliss |
|
|
Dit fenomeen wordt langetermijnpotentiatie genoemd en verwijst naar |
een toegenomen neuronale reactiviteit op een gewone (standaard) stimulatie in een pecifieke hersenregio naar aanleiding van een korte, intense elektrische stimulatie van enkele afferente neuronen die naar deze regio projecteren (Bliss |
|
|
In experimenteel onderzoek werden korte stroomstootjes toegebracht aan axonen van het perforerende baansysteem in de gyrus dentatus van de hippocampus. Terzelfdertijd werd de sterkte van de respons gemeten in areas die projectie van de gestimuleerde axonen ontvingen.
Nadat een stabiele basisrespons op de standaardstim u latie werd bekomen, werd een korte, hoogfrequente reeks van elektrische stimulatie toegebracht : een |
tetanus.
De hoogfrequente tetanische stimulatie werd dan stopgezet en de oorspronkelijke teststroomstoten werden hervat. De sterkte van de postsynaptische respons kan nu worden vergeleken met de originele basisrespons in functie van de tijd.
Vooreerst werd vastgesteld dat de sterkte van de excitatorische synaptische reacties van de hippocampale axonen aanzienlijk toeneemt onmiddellijk na de tetanische stimulatie. Deze toename neemt af met de tijd en keert terug naar de basisrespons. |
|
|
Vooreerst werd vastgesteld dat de sterkte van de excitatorische synaptische reacties van de hippocampale axonen aanzienlijk |
toeneemt onmiddellijk na de tetanische stimulatie. Deze toename neemt af met de tijd en keert terug naar de basisrespons. |
|
|
Deze kortetermijntoename wordt de … genoemd.
|
posttetanische potentiatie genoemd.
|
|
|
Een tweede vaststelling is dat na hoogfrequente tetanische stimulatie de postsynaptische reactiviteit in sommige hippocampale neuronen |
verhoogd blijft.
Toediening van een volgende 'standaard '-prikkel vertoonde een veel grotere respons dan voor de tetanische stimulatie.
|
|
|
Deze verhoogde responsiviteit kan uren, dagen en zelfs weken aanhouden en wordt daarom …. genoemd |
langeterrnijn potentiatie genoemd.
Langetermijnpotentiatie wordt geïnterpreteerd als een verhoging van synaptische efficiëntie.
|
|
|
Langetermijnpotentiatie wordt geïnterpreteerd als een verhoging van synaptische efficiëntie.
Gezien de sterkte van de standaardstimulus constant werd gehouden, kan elke verandering in reactiviteit worden beschouwd als een toename in |
synaptische effectiviteit.
Het fenomeen van langetermijnpotentiatie kan ook in andere regio's van de hersenen worden uitgelokt, maar werd vooral waargenomen in de formatio hippocampi (perforerend baansysteem en Schaffer colateraal baansysteem) , die zoals bekend verantwoordelijk wordt geacht voor de opslag van expliciete informatie.
|
|
|
Het fenomeen van langetermijnpotentiatie kan ook in andere regio's van de hersenen worden uitgelokt, maar werd vooral waargenomen in de |
formatio hippocampi (perforerend baansysteem en Schaffer colateraal baansysteem) , die zoals bekend verantwoordelijk wordt geacht voor de opslag van expliciete informatie.
|
|
|
Belangrijk is dat langetermijnpotentiatie kan worden geconditioneerd.
Wanneer een zwakke stimulus (die zelf niet in staat is een langetermijnpotentiatie op te wekken) wordt gekoppeld aan een sterke stimulus (die na enkele pogingen wel een langetermijnpoten tiatie kan uitlokken), dan zal de langeterrnijnpotentiatie |
ook optreden als reactie op de zwakkere stimulus.
Het is de associatie van de zwakke met de sterke prikkel die de langetermijnpotentiatie voor de zwakke stimulus mogelijk maakt.
|
|
|
Het is de associatie van de zwakke met de sterke prikkel die de langetermijnpotentiatie voor de zwakke stimulus mogelijk maakt.
Er is evenwel ook sprake van specificiteit. Andere zwakke prikkels die niet simultaan met de sterke stimulus werden gestimuleerd, zullen geen langeterrnijn potentiatie uitlokken (Brown, Chapman, Kairiss & Keenan, 1988), en de langetermijnpotentiatie geldt enkel voor die synapsen die door de stimulus werden geactiveerd .
Deze fenomenen zijn sterk vergelijkbaar met |
klassieke conditionering .
Een belangrijke voorwaarde voor het ontstaan van langeter rnijnpotentiatie blijkt een voldoende activatie (depolarisatie) van de postsynaptische cel.
Langeterrnijnpotentiatie vereist bijgevolg een simultane activatie van postsynaptische en presynaptische neuronen. Deze vaststelling is meteen het eerste bewijs voor Hebbs voorstel dat vooraleer leren kan optreden, zowel de presynaptische als de postsynaptische neuronen terzelfder tijd geactiveerd moeten zijn. |
|
|
Een belangrijke voorwaarde voor het ontstaan van langeter rnijnpotentiatie blijkt |
een voldoende activatie (depolarisatie) van de postsynaptische cel.
Langeterrnijnpotentiatie vereist bijgevolg een simultane activatie van postsynaptische en presynaptische neuronen. Deze vaststelling is meteen het eerste bewijs voor Hebbs voorstel dat vooraleer leren kan optreden, zowel de presynaptische als de postsynaptische neuronen terzelfder tijd geactiveerd moeten zijn. |
|
|
Langeterrnijnpotentiatie vereist bijgevolg een |
simultane activatie van postsynaptische en presynaptische neuronen.
Deze vaststelling is meteen het eerste bewijs voor Hebbs voorstel dat vooraleer leren kan optreden, zowel de presynaptische als de postsynaptische neuronen terzelfder tijd geactiveerd moeten zijn. |
|
|
Tot voor kort heerste het algemeen idee dat ionkanalen van neuronen (verantwoordelijk voor de overdracht van de prikkel) ofwel chemisch, ofwel elektrisch werden gereguleerd.
Tegenwoordig erkent men het bestaan van sommige synapsen in de gyrus dentatus waarbij de overdracht van ionen doorheen het membraan wordt gereguleerd door beide mechanismen. Deze ongewone ionkanalen maken deel uit van een complex apparaat, de
|
NMDA-receptor in het neuronaal membraan.
De meeste excitatorische synapsen van het hippocampaal systeem gebruiken het excitatorische aminozuur glutamaat als neurotransmitter. Afhankelijk van de eigenschap pen van de postsynaptische receptormolecule waarmee ze zich bindt, kan deze enkele neurotransmitter evenwel zeer verschillende effecten hebben in verschillende synapsen. Postsynaptische receptoren worden gecategoriseerd door de agonisten waarmee ze zich binden. |
|
|
De meeste excitatorische synapsen van het hippocampaal systeem gebruiken ………. als neurotransmitter. |
het excitatorische aminozuur glutamaat als neurotransmitter.
Afhankelijk van de eigenschappen van de postsynaptische receptormolecule waarmee ze zich bindt, kan deze enkele neurotransmitter evenwel zeer verschillende effecten hebben in verschillende synapsen. Postsynaptische receptoren worden gecategoriseerd door de agonisten waarmee ze zich binden. |
|
|
Afhankelijk van de eigenschappen van de postsynaptische receptormolecule waarmee ze zich bindt, kan deze enkele neurotransmitter |
evenwel zeer verschillende effecten hebben in verschillende synapsen.
|
|
|
Postsynaptische receptoren worden gecategoriseerd door de agonisten waarmee ze zich binden. Het belangrijkste onderscheid wordt gemaakt tussen receptoren die geactiveerd worden door
|
-N-methyl-D-a partaat (NMDA) en -receptoren die zich niet met deze stof verbinden.
|
|
|
Bij de niet-NMDA-receptoren gedraagt glutamaat zich als |
een perfect normale excitatorische neurotransmitter en induceert het een excitatorisch postsynaptisch potentiaal dat de postsynaptische cel tot vuren zal aansporen.
Wanneer de niet-NMDA-receptoren van het hippocampaal complex experimenteel worden geblokkeerd, verdwijnen ook alle excitatorisch postsynaptische potentialen .
|
|
|
Wanneer de niet-NMDA-receptoren van het hippocampaal complex experimenteel worden geblokkeerd, verdwijnen ook alle excitatorisch postsynaptische potentialen .
Omgekeerd heeft het blokkeren van alle NMDA-receptoren slechts een minimaal effect op de postsynaptische excitatie, maar stellen we vast dat |
elk spoor van langetermijnpotentiatie is verdwenen.
|
|
|
Het merkwaardige aan de NMDA-receptoren is dat zij |
twee afzonderlijke signalen moeten ontvangen alvoren ze geactiveerd kunnen worden .
Niet alleen dient de neurotransmitter glutamaat aanwezig te zijn , het postsynaptische membraan waarin de NMDA-receptor is gelegen, moet eveneen voldoende gedepolariseerd zijn , zoals door onmiddellijk voorafgaande excitatori che postsynaptische potentialen.
|
|
|
Het blijkt dat de langetermijnpotentiatie al volgt wordt geproduceerd (Figuur 4.13.).
Onder normale omstandigheden domineren de niet-NMDA-receptoren omdat de NMDA-receptoren geblokkeerd worden door een magnesiumion (Mg2+) en derhalve inactief zijn. De NMDA-receptoren kunnen we het best be chouwen als calciu mionkanalen. Door de magnesi u mblokkade blijft het calcium buiten de cel. Wa nneer de niet-NMDA-receptoren zich binden met de excitatorische tran mitter glutamaat, worden normale excitatorische postsynaptische potentialen geproduceerd. |
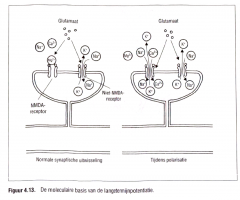
|
|
|
Onder normale omstandigheden domineren de niet-NMDA-receptoren omdat de NMDA-receptoren geblokkeerd worden door een magnesiumion (Mg2+) en derhalve inactief zijn.
De NMDA-receptoren kunnen we het best beschouwen als |
calciumionkanalen. Door de magnesiumblokkade blijft het calcium buiten de cel.
Wanneer de niet-NMDA-receptoren zich binden met de excitatorische tran mitter glutamaat, worden normale excitatorische postsynaptische potentialen geproduceerd. |
|
|
Gezamenlijke depolarisatie van synaptische input vanuit verschillende niet-NMDA-receptoren heft de magnesium blokkade van de NMDA-receptor op en maakt reactie op hoogfrequente input mogelijk.
Wanneer dit kanaal niet meer wordt geblokkeerd door het magnesiumion én wanneer de NMDA-re ceptor zelf voldoende gedepolariseerd is door glutamaat, en alleen dan, kunnen |
de calciumionen (Ca2+) (en natriumionen (Na+)) de cel binnenstromen.
Het calcium blijkt fundamenteel voor het opwekken van een langetermijnpotentiatie. |
|
|
Veronderstel nu even dat twee nabijgelegen (axonale) excitatorische synapsen verbinding maken met één (dendritische) postsynaps (Figuur 4.14.).
Ter hoogte van de excitatorische synaps A zijn de postsynaptische receptoren van het niet-NMDA-type, en ter hoogte van synaps B zijn ze van het NMDA-type.
|
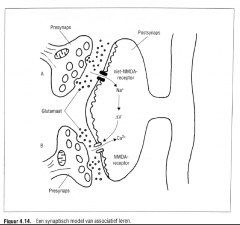
|
|
|
Wanneer de synapsen afzonderlijk actief zijn, zal het NMDA-kanaal |
geen calcium toelaten .
Als synaps A daarentegen vlak voor synaps B actief is, wordt via de niet-NMDA-receptor ook het NMDA-kanaal gedepolariseerd, verdwijnt de magnesiumblokkade van de NMDA-receptor, en zorgt de eveneens noodzakelijke activatie van het glutamaat ter hoogte van de NMDA-receptor dat het calcium via het NMDA-kanaal de postsynaptische cel kan binnenstromen.
|
|
|
Wanneer synaps B actief is voor synaps A, blijft het calciumkanaal van de NMDA-re ceptor geblokkeerd door het magnesium.
Merk op dat het receptorcomplex twee criteria van associatief Ieren vervult, namelijk |
twee aparte signalen (gebeurtenissen) hebben plaats binnen een specifieke temporele orde (cfr. klassieke conditionering).
Het feit dat het postsynaptisch neuron reeds geactiveerd (gedepolariseerd) moet zijn om langeter mijnpotentiatie mogelijk te maken komt goed overeen met het principe van de Hebbiaanse synaps. |
|
|
De aanwezigheid van calcium is noodzakelijk voor
|
het ontstaan van een langetermijn potentiatie.
Calcium faciliteert synaptische activatie door de activiteit van het calciumaf hankelijke proteïnekinase CAM kinase II.
|
|
|
Calcium faciliteert synaptische activatie door de activiteit van het calciumaf hankelijke proteïnekinase CAM kinase II.
Genetisch gemanipuleerde muizen met een CAM kinase II-deficiëntie vertonen een |
verminderde capaciteit voor langetermijnpoten tiatie en presteren slecht op de voornoemde waterdoolhoftest (Silva , Stevens, Tonegawa
De werking van CAM kinase II is echter ingewikkeld en het precieze substraat dat verantwoordelijk is voor de langetermijnpotentiatie en leerprocessen, is vooralsnog onduidelijk.
|
|
|
Hoewel aangenomen wordt dat langetermijnpotentiatie het resultaat is van een verhoogde synaptische efficiëntie, is het onderliggende mechanisme van de synaptische veranderingen |
nog niet gekend. Zie Kandel, Schwartz en Jessell (1995) voor een meer gedetailleerd overzicht van de cellulaire mechanismen van leren en geheugen. |