![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
78 Cards in this Set
- Front
- Back
|
1.9 Neuronale prikkeloverdracht
De signalen die de neuronen gebruiken om met elkaar te communiceren zijn elektrochemische signalen.
Een actiepotentiaal is een plotselinge verandering van elektrische polariteit tussen de binnen- en buiten kant van een axon die word t veroorzaakt als |
een reactie op stimulatie van de zenuwcel (door bijvoorbeeld andere zenuwcellen of, in het geval van zintuiglijke neuronen , door zintuiglijke prikkels).
Actiepotentialen dienen om informatie vanuit het cellichaam doorheen het axon naar de eindplaatjes van het neuron te verzenden.
|
|
|
Deze alles-of-nietspotentialen voorzien in informatieoverdracht over grotere afstanden binnen één enkel neuron.
Voor dit deel van de prikkeloverdracht wordt gebruik gemaakt van elektrofysiologische mechanismen. Uiteindelijk bereikt het actie potentiaal de knopvormige eindplaatjes van het axon.
Dit eindknopje is gevuld met |
kleine blaasjes of synaptische vesikels die de neurotransmitters bevatten.
Door de elektrofysio logische stimulatie komt de transmitterstof vrij in de ruimte tussen twee cellen die met elkaar contact hebben gezocht, de synaptische spleet.
|
|
|
Door het vrijkomen van neurotransmitters in deze synaptische spleet prikkelt het neuron de receptoren die gelegen zijn op het celmembraan van het postsynaptisch neuron.
Het effect van de neurotransmitter op deze postsynaptische cel kan zowel |
exciterend als inhiberend zijn .
Wanneer transmitter en receptor excitatorisch zijn, kunnen ze het celmembraan van de postsynap tische cel depolariseren en aldus de kans verhogen dat deze cel zal 'vuren' , dat wil zeggen een nieuw actiepotentiaal doen ontstaan.
Wanneer daarentegen de transmitter en receptor inhibitorisch van aard zijn, kunnen ze het membraan hyperpolariseren en aldus de kans verkleinen dat deze cel zal gaan vuren.
De werking van de neurotransmitters ter hoogte van de synaptische spleet zorgt voor de prikkeloverdracht tussen verschillende neuronen en berust vooral op een chemische basis . |
|
|
De werking van de neurotransmitters ter hoogte van de synaptische spleet zorgt voor de prikkeloverdracht tussen verschillende neuronen en berust vooral op |
een chemische basis . |
|
|
Het verschil in voltage tussen de binnenkant (intracellulaire vloeistof) en de buitenkant (extracellulaire vloeistof) van het celmembraan resulteert in |
een elektrisch potentiaalverschil dat met behulp van micro-elektroden kan worden gemeten.
Het rustpotentiaal van een zenuwcel is ongeveer -70 millivolt (mV ; een millivolt is één duizendste van een volt) met een negatieve lading binnenin de cel.
|
|
|
De term rustpotentiaal is enigszins misleidend omdat het neuron een flink deel van zijn energie moet aanwenden om deze toestand te handhaven.
De negatieve intracellulaire lading wordt veroorzaakt door relatieve verschillen in concentratie van positief en negatief geladen deeltjes (ionen) aan weerszijden van het celmembraan.
Ionen zijn |
atomen die één elektron te veel of te weinig hebben, wat resulteert in een discrepantie tussen de positief geladen protonen en de negatief geladen elektronen in het atoom.
|
|
|
Atomen met een extra elektron worden anionen genoemd en zijn negatief geladen.
Atomen met een elektron te weinig worden kationen genoemd en zijn (door het extra proton) positief geladen.
Anionen en kationen hebben derhalve een tegengestelde lading. De geladen deeltjes reageren als |
kleine magneetjes waarbij tegen
Wanneer anionen en kationen worden gescheiden, bestaat er een kracht die beide ionen naar elkaar trekt.
Ook wanneer er een barrière (zoals een celmembraan) tussen de twee ladingen aanwezig is, blijft deze kracht bestaan .
|
|
|
Die kracht, de arbeid noodzakelijk om beide ladingen naar elkaar toe te brengen , wordt het voltage of het potentiaal verschil genoemd.
Wanneer de barrière wordt verwijderd, bewegen de ladingen |
naar elkaar toe. Deze beweging noemen we stroom.
Stroom is de mate waarmee de ladingen bewegen. Zowel het cytoplasma (de intracellulaire vloeistof) als de extracelluJaire vloeistof is rijk aan ionen.
|
|
|
De belangrijkste ionen die een rol spelen in het ontstaan van een potentiaalverschil, zijn
n k c o
|
-de positief geladen ionen Natrium (Na+) en -Kalium (K+) en -de negatief geladen Chloor (Cl-) en -organische anionen (An-).
|
|
|
Indien de ionen zich vrij in een oplossing zouden bevinden, gaan ze zich evenredig over deze oplossing verdelen.
Ze gehoorzamen daarbij aan twee krachten:
e c
|
-elektrische lading (positief versus negatief) en -concentratie (een stof zal zich homogeen over een oplossing verspreiden).
Het potentiaalverschil wordt vooral bewerkstelligd door het celmembraan dat de intracellulaire ruimte afscheidt van de extracellulaire ruimte.
De concentraties van verschillende ionen in de extracellulaire en intracellulaire vloeistoffen zijn namelijk aanzienlijk verschillend. |
|
|
De verschillende ionconcentraties binnen en buiten de cel worden door het celmembraan op verschillende manieren bewerkstelligd .
Eerst en vooral bevat de cel zelf een grote concentratie aan negatief geladen organische anionen An· die gewoon te groot zijn om door het celmembraan naar buiten te dringen en dus in de intracellulaire ruimte zijn opgesloten.
Ten tweede is het celmembraan op een gedifferentieerde manier semi doorlaatbaar of emi-permeabel voor bepaalde ionen.
De kleinere K+-ionen raken vlotter door het celmembraan dan de grotere Na+-ionen. Het celmembraan bevat afzonderlijke K+- en Na+-kanalen (gespecialiseerde proteïnestructuren) die afhankelijk van de omstan digheden hun doorlaatbaarheid voor deze ionen kunnen aanpassen.
In rusttoestand laat het celmembraan enkel |
K+-ïonen toe terwijl de in- en uitgaande beweging van alle andere molecuultypes wordt verhinderd.
Doorheen kleine poriën of kaliumkanalen kunnen de kaliumionen zich vrij van binnen naar buiten of van buiten naar binnen de cel bewegen .
|
|
|
Aangezien er veel meer kaliumionen in de intracellulaire vloeistof zijn dan in de extracellulaire vloeistof , zullen de kaliumionen geneigd zijn de intracellulaire vloeistof via het celmem braan te verlaten naar de extracellulaire vloeistof.
Het concentratieverschil van kaliumionen tussen intra- en extracellulaire ruimte creëert immers |
een concentratie gradiënt door het membraan.
Wanneer de kaliumionen zich vrij doorheen bet membraan mogen bewegen , zal de resultante beweging op basis van de concentratiegradiënt er een zijn naar een gebied van lage concentratie. |
|
|
Omdat de kaliumionen geladen deeltjes zijn en ook het enige type van ionen die vrij doorheen het celmembraan kunnen bewegen, gaat de beweging van de positief geladen kaliumionen van binnen naar buiten de cel een elektrisch onevenwicht of lading veroorzaken.
De uitwaartse diffusie van kaliumionen volgens de concentratiegradiënt resulteert in |
een verlies van positieve ionen aan de binnenkant van de cel en een teveel aan positieve ionen aan de buitenkant waardoor een membraanpotentiaal ontstaat.
De binnenkant van de cel wordt negatiever geladen dan de buitenkant (denk aan de negatief geladen grote organische anionen die in de cel zitten opgesloten). Het ontstaan van dit membraanpotentiaal heeft ook gevolgen voor de beweging van de kaliumionen.
|
|
|
Wanneer de positief geladen kaliumionen blijven diffunderen vanuit de cel, wordt het membraanpotentiaal steeds groter. Positief geladen deeltjes worden echter afgestoten door andere positieve ladingen en aangetrokken door negatieve ladingen.
Wanneer het membraanpotentiaal dus groter wordt, ontstaat er eveneens een |
elektrische kracht die de uitwaartse migratie van de kaliumionen gaat tegenwerken en de kaliumionen terug in de cel trekt.
Uiteindelijk ontstaat er een evenwicht waarbij het membraanpotentiaal voldoende sterk is ten aanzien van de concentratiegradiënt. Voor elk kaliumion dat uit de cel diffundeert door de kaliumkanalen van het membraan, wordt een ander door de (negatieve) intracellulaire elektrische lading door het mem braan terug in de cel getrokken.
Er is een elektrochemisch equilibrium ontstaan en de resulterende beweging van kaliurnionen is nul. |
|
|
1.9.2 De natrium-kaliumpomp
Hodgkin en Huxley (Nobelprijs in 1963) verfijnden Bernsteins hypothese van de kalium ionen en toonden aan dat een ander ion, natrium, ook een aandeel heeft in het bepalen van het rustpotentiaal .
De concentratie van natrium is ongeveer negen keer groter buiten de cel dan binnen de cel. Natriumionen zijn positief geladen en gezien het rustpotentiaal binnen de cel -70 mV is, bevinden de natriumionen zich in een elektrochemisch onevenwicht.
Zowel qua concentratiegradiënt als qua lading willen de natriumionen zich in de cel begeven, maar worden ze tegengehouden door
|
het celmembraan.
Toch is het celmembraan in geringe mate ook doorlaatbaar voor natrium. De natriumdoorlaatbaarheid is ongeveer een honderdste van de doorlaatbaarheid voor kalium wanneer het membraan in rust is.
|
|
|
De kleine inwaartse stroom van natriumionen wordt veroorzaakt door de concentratiegradiënt van natrium (hogere concentratie buiten de cel dan binnen de cel) en de elektrische gradiënt die wordt veroorzaakt door het (wegstromende) kalium. De positief geladen natriumionen worden aangetrokken door de negatief geladen intracellulaire vloeistof.
Ondanks de weliswaar beperkte doorlaatbaarheid voor natrium zou een inwaartse stroom van natrium ionen na verloop van tijd bet door de kalium ionen ontstane potentiaalverschil kunnen opheffen.
Onder invloed van hun verschillende concentratie gradiënten zouden natriumionen de intracellulaire vloeistof blijven |
binnendruppelen en kaliumionen zouden de intracellulaire vloeistof verlaten. Deze in- en uitstroom van ionen is niet erg groot maar wel gestaag.
Na verloop van tijd zou de inwendige concentratie van natrium steeds groter worden en de inwendige concentratie van kalium steeds kleiner. Het verschil inelektrische lading tussen de binnenkant en de buitenkant van het membraan zou verdwijnen. |
|
|
Het mechanisme dat deze gang van zaken echter verhindert, noemt men de natrium-kaliumpomp. Het celmembraan beschikt over een metabolisch pompsysteem dat actief het intracellulair natrium uit de cel drijft en de ontsnapte extracellulaire kaliumionen terug in de cel brengt.
Onderzoek wijst uit dat |
een drietal natriumionen worden uitgewisseld tegen twee kaliumionen .
Hoewel de kaliumionen (met relatief vrije mem braandoorgang) vrijwel onmiddellijk de cel terug zullen verlaten, geldt dit veel minder voor de natriumionen gezien het membraan minder doorlaatbaar is voor Na+ dan voor K+.
|
|
|
Gelet op deze pompfunctie ter hoogte van het celmembraan en gezien de overwegend negatieve lading van de intracellulaire vloeistof, stapelt het Na+ zich op aan de buitenkant van het celmembraan. De uitwaaitse beweging van natrium (tegen concentratiegradiënt en elektrisch potentiaal in) wordt veroorzaakt door een actief membraan systeem dat energie verbruikt. Ongeveer 40% van het energieverbruik van de cel wordt besteed aan deze natrium-kaliumpomp.
|
eerder passieve wijze op chemische en elektrische gradiënten en dragen n iet bij tot het membraan potentiaal dat door de actieve K+- en Na+-ionen wordt bewerk telligd. Gezien het membraan doorlaatbaar is voor de negatief geladen chloorionen , volgt het chloor het natrium dat door de natrium-kaliu mpomp uit de cel in de po itief geladen extracellulaire vloeistof wordt gepompt.
De pomp is dus ook verantwoordelijk voor het verwijderen van chloorionen uit het cytoplasma , hoewel dit en indirect effect is. |
|
|
De pomp is dus ook verantwoordelijk voor het verwijderen van chloorionen uit het cytoplasma , hoewel dit |
een indirect effect is. |
|
|
1.9.3 Het actiepotentiaal
Wanneer een zenuwcel wordt gestimuleerd (bijvoorbeeld door het axon van een andere cel), verandert haar elektrische polariteit.
Het celmembraan reageert op stimuli door zijn doorlaatbaarheid voor ionen te veranderen.
Afhankelijk van het soort van stimulatie kan het potentiaalverschil groter of kleiner worden. Wanneer de stimulatie leidt tot een daling van het potentiaalverschil , spreken we van een |
depolarisatie van het membraan.
Wanneer de stimulatie een verhoging van het potentiaalverschil veroorzaakt , spreken we van een hyperpolarisatie van het membraan.
|
|
|
Depolarisatie wordt veroorzaakt door een verhoogde inwaartse stroom van natriumionen.
Hyperpolarisatie wordt waarschijnlijk veroorzaakt door |
een verhoogde uitstroom van kalium.ionen of een verhoogde instroom van chloor ionen. |
|
|
Hoe groter de depolarisatie , hoe meer kanalen er worden geopend. Omdat natrium een positief geladen ion is, dat zich in hoge concentraties aan de buitenkant van het celmembraan bevindt , resulteert de opening van de natriumkanalen in |
een inwaartse beweging van de positieve natriumionen en dus in een verdere depolarisatie van het rustpotentiaal.
|
|
|
Weinig prikkeling van de cel resulteert in een kleine depolarisatie waarbij slechts enkele natrium.kanalen worden geopend .
Wanneer de daar door veroorzaakte instroom van natrium minder is dan de normale uitstroom van kalium , gaat de depolarisatie wegebben en keert de cel terug naar |
zijn rustpotentiaal.
In dit geval wordt er geen actiepotentiaal uitgelokt.
Wanneer de depolarisatie echter zo groot is dat meer natriumionen bet neuron binnenstromen dan er kaliumionen de cel uitstromen , wordt een bepaalde grenswaarde overschreden (-50 mV) en wordt het membraan (tijdelijk) natriumdoorlaatbaar waarbij de natriumionen massaal de cel binnenstormen en het membraanpotentiaal van negatief naar positief (+40 mV) ombuigen. |
|
|
Er zijn twee factoren die ervoor zorgen dat het actiepotentiaal wordt afgebroken en die bet membraan terug in de rusttoestand brengen.
|
-Ten eerste is de opening van de natriumkanalen een zeer kortstondig proces. Na de opening worden ze onmiddellijk gesloten en worden ze inactief ; ze kunnen gedurende verschillende milliseconden niet meer worden geopend zodat de invoer van nieuwe natriumionen wordt verhinderd.
-Ten tweede verhoogt het celmembraan zijn doorlaatbaarheid voor kalium . De verhoogde membraanpermeabiliteit voor kalium versnelt de terugkeer naar het rustpotentiaal. |
|
|
De inactivatie van de natriumkanalen en de toegenomen doorlaatbaarheid van bet membraan voor kalium zorgt voor een tijdelijke onmogelijkheid om een nieuw actiepotentiaal te vormen : de |
absolute refractaire periode . In de meeste neuronen kan er na een actiepo tentiaal gedurende ongeveer 1 milliseconde geen nieuw actiepotentiaal worden geproduceerd. De doorlaatbaarheid voor kalium blijft ook wat langer gehandhaafd en dit resulteert in een tijdelijke hyperpolarisatie van het neuron (dat wil zeggen een verhoging van het membraanpotentiaal tot -75mV) en dus in een verhoging van de grenswaarde waardoor
|
|
|
De instroom van natrium moet de uitstroom van kalium inuners overtreffen om over de grenswaarde te gaan en een neuw actiepotentiaal uit te lokken .
Deze periode van verhoogde grenswaarde na de absolute refractaire periode wordt de |
relatieve refractaire periode genoemd.
Tijdens de relatieve refractaire periode is er dus een sterker dan normale input nodig om een nieuw actiepotentiaal uit te lokken. |
|
|
Actiepotentiaal voorstelling: |
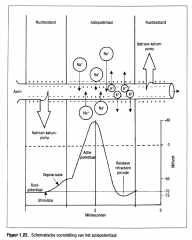
|
|
|
Actiepotentialen worden geproduceerd door veranderingen in de voltagegevoelige natrium- en kaliumkanalen van het axonaal membraan maar het is de richting waarin deze ladingsveranderingen verlopen die het actiepotentiaal over het axon doet bewegen.
Dit gebeurt doordat |
de elektrische ladingen die worden geproduceerd door de bewegingen van de ionen, zich over het axon verspreiden en aanpalende gebieden van het membraan depolariseren.
|
|
|
De depolariserende effecten van het actiepotentiaal kunnen zich in aanzienlijke mate over het axon verspreiden en deze verspreide depolarisatie lokt andere actiepotentialen uit waarbij de beweging van het celmembraan naar de eindplaatjes is gericht.
Op deze manier reist het actiepotentiaal langsheen het axon als een golf van |
elektrochemische energie.
Hoewel het membraan wordt gedepolariseerd langs beide zijden van het actiepotentiaal, beweegt het potentiaal zich enkel in voorwaartse richting.
|
|
|
De zenuwimpuls kan zich niet verplaatsen naar de regio van het membraan van waar het is gekomen omdat dit weefsel zich in een refractaire periode bevindt en er gedurende korte tijd geen nieuw actiepotentiaal kan worden gevormd.
De impuls beweegt zich in |
één richting van axonheuvel naar eindplaatjes voort, ook al spreidt de depolarisatie zich in beide richtingen rond de actieve site.
|
|
|
Wanneer deze spreiding klein is, zal het actiepotentiaal |
enkel het membraan in zijn onmiddellijke omgeving beïnvloeden en zal de snelheid van de golf traag zijn.
Wanneer het depolariserend effect van de spike echter breder is, zal het actiepotentiaal zich sneller over het celmembraan bewegen.
|
|
|
De snelheid van de zenuwimpul is dus afhankelijk van de afstand waarmee het actiepotentiaal een tegen de grenswaarde depolariserend effect heeft.
Anders gezegd, |
een hogere passieve spreiding van de depolariserende stroom levert een hogere gelei dingssnelheid.
|
|
|
Grotere vezels hebben een hogere geleidings snelheid dan kleinere omdat de depolariserende effecten van het actiepotentiaal zich verder over het axon kunnen verspreiden.
Een andere manier om de geleidingssnelheid van de neuronen te verhogen is door |
het axon te isoleren (zoal een elektrische draad wordt geïsoleerd door middel van niet-geleidend plastic) zodat de effectiviteit van het voortsnellende actiepotentiaal kan worden verhoogd.
|
|
|
Het isolatiemateriaal van het zenuwstelsel betaat uit het reeds genoemde myeline dat door de gliale cellen (oligodendrocyten in het centraal en Schwann cellen in het perifeer zenuwstelsel) wordt geproduceerd en zich als een schede rond het axon wikkelt.
Vrijwel alle belangrijke zenuwbanen binnen het menselijke zenuwstelsel zijn gemyeliniseerd. Myelinisatie laat toe dat |
complexe communicatie tussen de verschillende regio's van het zenuwstelsel zeer nel kunnen verlopen. |
|
|
Het belangrijkste effect van myelinisatie op de voortbeweging van de zenuwimpuls is dat ze het contact verhindert tussen het axonmembraan en de extracellulaire vloeistof.
Op de gemyelinieerde regio zelf kan dus geen ionuitwisseling plaat vinden en kunnen ook geen |
actiepotentialen worden uitgelokt.
Actiepotentialen hebben enkel plaats ter hoogte van de blootgestelde uitsparingen tussen de egmenten van de myelineschede.
|
|
|
Deze uitsparingen worden .... genoemd |
de knopen van Ranvier genoemd.
Myelinisatie versnelt het actiepotentiaal door de ionuitwisselingen over het celmembraan in de gemyeliniseerde regio te elimineren en te beperken tot de uitsparingen tussen de myeline segmenten .
|
|
|
Door de ionuitwisselingen enkel op deze knopen toe te laten wordt de stroom die zich rond het actiepotentiaal verpreidt, sneller geconcentreerd naar een volgende plaats op het axon.
Het depolariserend effect van een actiepotentiaal dat wordt opgebouwd in een knoop van Ranvier aan één kant van het myelinesegment , verspreidt zich langs |
het axon naar de volgende knoop waar een nieuw actiepotentiaal wordt geproduceerd.
Ter hoogte van elke knoop wordt een nieuw actiepotentiaal uitgelokt.
|
|
|
Tussen knopen zijn er geen actiepotentialen maar enkel een spreiding van depolariseren de stroom van één knoop naar de volgende.
Dit effect wordt .... genoemd |
saltatorische conductie genoemd .
Omdat actiepotentialen enkel en alleen uitgelokt worden ter hoogte van de knopen van Ranvier en niet onder de myelineschede , kan men hier ook enige specialisatie van het axonmem braan verwachten.
Het celmembraan ter hoogte van een knoop is uitzonderlijk rijk aan zowel kalium- als natriumkanaaltjes . |
|
|
Naast het verhogen van de snelheid van de zenuwgeleiding veroorzaakt myeline ook |
een vermindering van de metabolische inspanning van de impulstrans missie.
Wanneer de productie van actiepotentialen beperkt blijft tot de knopen van Ranvier, betekent dit dat ongeveer 1% van het axonmembraan bij het proces betrokken is.
Het aantal ionbewegingen wordt daardoor sterk gereduceerd en de noodzakelijke arbeid van de energieverbruikende kalium-natriumpomp wordt bijgevolg ook veel kleiner. |
|
|
Wanneer de productie van actiepotentialen beperkt blijft tot de knopen van Ranvier, betekent dit dat |
ongeveer 1% van het axonmembraan bij het proces betrokken is.
Het aantal ionbewegingen wordt daardoor sterk gereduceerd en de noodzakelijke arbeid van de energieverbruikende kalium-natriumpomp wordt bijgevolg ook veel kleiner. |
|
|
Het aantal ionbewegingen wordt daardoor sterk gereduceerd en de noodzakelijke arbeid van de energieverbruikende kalium-natriumpomp wordt bijgevolg ook |
veel kleiner. |
|
|
Het actiepotentiaaI wordt ook wel een zen uwimpuls of spike genoemd vanwege |
zijn scherpe, bijna explosieve karakter.
Om een meer plastische beschrijving te geven kan men zeggen dat het axon (het neuron) 'vuurt'.
|
|
|
Gewoonlijk wordt bet actiepotentiaal uitgelokt ter hoogte van de axonheuvel in de overgang tussen cellichaam en axon.
Van daaruit verspreidt het zich over het hele axon zoals een vuurtje dat langs een lont loopt. Een actiepotentiaal heeft een aantal eigenaardige kenmerken. |
-Eerst en vooral is het geen graduele maar een alles-of-niets gebeurtenis : ofwel is er een actiepotentiaal ofwel is er geen. -Ten tweede zijn alle actiepotentialen van ongeveer dezelfde grootte en bestaan er geen 'zwakke' of 'sterke' actiepotentialen. Actiepotentialen communiceren informatie door hun aanwezigheid, niet door veranderingen in vorm of grootte. -Ten derde worden de actiepotentialen voortgestuwd door het axon zelf. Het signaal moet voortdurend worden herschapen door het mem braan van het axon om het van de axonheuvel naar de eindplaatjes te brengen.
|
|
|
Tot hiertoe bleven we erg vaag over de oorsprong van het actiepotentiaal.
We schreven dat de cel wordt 'geprikkeld' en dat deze prikkeling leidde tot een 'depolarisatie ' die, indien sterk genoeg, aanleiding gaf tot een actiepotentiaal dat vanaf de axonheuvel op het cellichaam over de hele lengte van het axon naar de axonale eindplaatjes loopt.
In feite vormen de dendrieten de sensibele tentakels van het neuron . Ook de dendrieten beschikken over een gelijkaardig celmembraan en rustpotentiaal en ondergaan potentiaalveranderingen wanneer ze worden geprikkeld.
Dendrieten produceren echter geen |
actiepoten tialen.
Wanneer een dendriet wordt gestimuleerd, staan de gemeten potentiaalverande ringen in verhouding met de intensiteit van de ontvangen stimulatie.
De potentiaalveran dering spreidt zich over de dendriet en wordt zwakker naarmate het zich verder van de stimulatiebron verwijdert.
|
|
|
De dendritische potentiaalveranderingen worden graduele potentialen genoemd en kunnen afhankelijk van het soort stimulatie een afname (depolarisatie) of toename (hyperpolarisatie) in het membraanpotentiaal veroorzaken.
Wanneer een dendriet op twee naburige plaatsen wordt gestimuleerd en wanneer beide prikkels van gelijke aard zijn (bijvoorbeeld allebei depolariserend) , dan worden beide effecten |
bij elkaar opgeteld en zal de potentiaalverandering groter zijn dan wanneer maar één prikkel werd ontvangen.
Wanneer beide stimuli te ver van elkaar ontstaan, dan zullen hun effecten uitgedoofd zijn alvorens ze elkaar kunnen bereiken.
|
|
|
Op gemiddelde afstand van elkaar zal enkel de resterende invloed van de reeds gedeeltelijk uitgedoofde prikkels kunnen worden opgeteld, en wanneer de stimuli van tegengestelde soort zijn (de ene depolarise rend en de ander hyperpolariserend) , |
heffen ze elkaar gewoon op.
|
|
|
Dit mechanisme waarbij naburige graduele potentialen met elkaar worden opgeteld of afgetrokken, wordt |
spatiële sommatie genoemd.
Temporele sommatie volgt een gelijkaardig principe. De graduele potentiaal van een gestimuleerde dendriet dooft langzaam uit.
|
|
|
Wanneer een tweede stimulus onmiddellijk na de eerste stimulus volgt, zullen beide effecten worden opgeteld (weer op voorwaarde dat beide prikkels van dezelfde soort zijn, anders heffen ze elkaar natuurlijk op).
Hoe meer tijd er tussen beide stimuli ligt, hoe kleiner het gezamenlijk effect. De sterkte van een graduele potentiaal zal dus worden bepaald door |
de sterkte (en de soort), de nabijheid en het tijdsinterval tussen de ontvangen stimuli.
Wanneer het dendritisch systeem dermate wordt geprikkeld dat er een depolarisatie tot
Wanneer de graduele potentialen niet voldoende sterk zijn om de grenswaarde van -50 mV te bereiken, zal er geen actiepotentiaal ontstaan. |
|
|
1.9.6 Synaptische activiteit
Het is via deze synapsen dat de ene cel de activiteit van een andere cel kan beïnvloeden door middel van chemische stoffen die worden losgelaten wanneer het neuron een actiepotentiaal afvuurt.
In de synaps wordt het celmembraan van het 'vurende ' neuron het |
presynaptisch membraan genoemd .
Het membraan van de ontvangende cel wordt het postsynaptisch membraan genoemd.
|
|
|
De ruimte tussen presynaptisch en postsynaptisch membraan noemen we de synaptische spleet .
Deze spleet is gevuld met |
een chemisch complexe vloeistof.
De synaptische spleet bevat ook kleine filamentjes of draadjes die de presynaptische en postsy naptische membranen dicht bij elkaar houden : het synaptische web.
|
|
|
Zowel dendrieten als axonen hebben presynaptische eindplaatjes en zowel axonen, dendrieten als het celmembraan zelf kunnen een postsynaptisch element zijn. Vrijwel elke denkbare combinatie van microanatomische synaps vorming is mogelijk.
Verpakt in een eigen vesikel worden de neurotransmitters van uit het cellichaam getransporteerd naar en bewaard in |
de eindplaatjes.
De eindplaatjes zijn rijk aan mitochondriën, wat een anatomische indicatie is dat de biochemische mechanismen van het eindplaatje een grote nood hebben aan hoog energetisch materiaal.
|
|
|
In een chemische synaps veroorzaakt de aankomst van een zenuwimpuls (actiepotentiaal) ter hoogte van het presynapti sche eindplaatje van het axon het openen van enkele synaptische vesikels die een chemische substantie, een neurotransmitter of neuromodulator loslaten.
Het proces waarbij de vesikels in het eindplaatje versmelten met het celmembraan en hun inhoud in de synaptische spleet loslaten , wordt .... genoemd. |
exocytose genoemd.
Nadien sluit het vesikel zich opnieuw en gaat het terug naar het inwendige van het eindplaatje waar het opnieuw met neurotransmitter wordt gevuld.
De neurotransmitter reist doorheen de synaptische spleet en valt op het celmembraan van de postsynaptische cel.
|
|
|
Het postsynaptisch membraan bevat een aantal gespecialiseerde proteïnen die dienstdoen als ontvangers voor de neurotransmitters.
De gespecialiseerde proteïnemoleculen liggen ingebed in bet postsynaptisch membraan en binden zich aan de extracellulaire zijde van het membraan in de synaptische spleet met een specifieke neurotransmittermolecule.
De neurotransmitter wordt vervolgens |
weggewassen en onschadelijk gemaakt door de extracellulaire vloeistof of voor hergebruik opnieuw in het presynaptisch membraan opgenomen.
|
|
|
Wanneer de receptor zich bindt met een neurotransrnittermolecule , gaat de ermee verbonden effector aan de binnenzijde van het membraan de postsynaptische reactie produceren.
Meestal gebeurt dit door |
het openen van ionkanalen waardoor een exciterend (depolariserend) of inhiberend (hyperpolarise rend) postsynaptisch potentiaal wordt bewerkstelligd.
Gezien de receptorkant in principe losstaat van de effectorzijde en er verschillende effectortypes bestaan, kan eenzelfde neurotransmitter verschillende fysiologische effecten hebben bij verschillende receptor effectorsamenstellingen.
|
|
|
Een receptor-effectorcomplex wordt dus niet alleen bepaald door het soort neurotransmitter waarmee het zich bindt (rol van de receptor), maar ook door de aard van de postsynaptische respons die het veroorzaakt (rol van de effector).
De meest gebruikelijke manier om neurotransmitters en receptor-effectorcomplexen te bestuderen is door |
het effect van verschillende chemische stoffen op de synaptische werking te evalueren.
Sommige stoffen zijn in staat zich met een receptor te binden en het effectorsysteem te activeren .
|
|
|
Deze stoffen gedragen zich dus al of ze een bepaalde neurotransmitter zijn en worden |
agonisten genoemd.
De drug LSD bijvoorbeeld is een agonist van de neurotransmitter serotonine.
|
|
|
Andere chemische stoffen, de antagonisten, verminderen of blokkeren de respons van een receptor -effectorcomplex op een neurotransmitter of zijn agonist.
Dit kan bijvoorbeeld door |
de receptorzijde te 'bezetten' zonder een reactie van de effector te veroorzaken en aldus het effect van de neurotransmit ter te blokkeren.
|
|
|
Net zoals kalium het rustpotentiaal domineert en natrium het actiepotentiaal bestuurt, regelt calcium het loslaten van een neurotransmitter in de synaptische spleet.
De hoeveelheid neurotransmitter die wordt lo gelaten in een synaps, is sterk gecorreleerd met |
de concentratie van calciumionen (Ca2+) in de extracellulaire vloeistof.
Vermoed wordt dat elk actiepotentiaal de opening van calcium electieve kanalen ter hoogte van het synaptisch membraan veroorzaakt en dat het calcium bet loslaten van de neurotransmitter door de vesikels in de synapti cbe spleet bewerkstelligt.
|
|
|
Een dergelijke proefopstelling leert dat |
de postsynaptische graduele potentialen onmiddellijk reageren op elke presynaptische prikkeling.
Bovendien blijken bepaalde presynapti che axonen een depolariserend effect op het postsynaptisch celmembraan te hebben terwijl andere een hyperpolariserend effect hebben. |
|
|
Depolariserende postsynaptische potentialen verhogen de kans dat het neuron zal vuren en worden daarom excitatorische postsynaptische potentialen (EPSP's) genoemd.
Hyperpolariserende postsynaptische potentialen maken het neuron minder ge voelig voor ontlading en worden |
inhibitorische postsynaptische potentialen (IPSP's) genoemd.
De graduele potentialen van de dendrieten worden veroorzaakt door de invloed van de neurotransmitters vanuit de eindplaatjes van presynaptische neuronen .
|
|
|
Duizenden eindplaatjes synapteren met de dendrieten en het cellichaam van een neuron en het resulterend gradueel potentiaal van de cel is het resultaat van al deze invloeden.
Het is de (spatiële en temporele) som van al deze prikkeling die bepaalt of |
het neuron zal vuren of niet. Wanneer de excitatorische potentialen overheersen en samen van voldoende sterkte zijn om de grenswaarde aan de axonheuvel te overschrijden, zal de cel een actiepotentiaal afvuren. Indien inhibitorische potentialen overheersen, zal het nemon niet vuren .
Het vuren van het neuron is een alles-of-niets mechanisme dat doorgaat zolang de grenswaar de ter hoogte van de axonheuvel behouden blijft.
Het elektrofysiologische actiepotentiaal zorgt op zijn beurt via chemische mechanismen ter hoogte van de eindplaatjes voor de overdracht van de prikkel op het volgende postsynaptische neuron. |
|
|
Om een chemische stof als een neurotransmitter te kunnen identificeren dient deze stof aan een aantal voorwaarden te voldoen :
|
|
|
|
Op grond van deze criteria en rekening houdend met het feit dat de huidige technologie nog niet toereikend is om al deze criteria ten gronde te onderzoeken, wordt een tiental chemische stoffen als waarschijnlijke neurotransmitters beschouwd.
Naast de klassieke neurotransmitters bestaat er een dertigtal |
peptiden (ketens van aminozuren) die zich in de zenuwcellen bevinden en een rol spelen in de neurotransmissie.
Deze neuropeptiden worden ook wel neuromodulators genoemd. Klassieke neurotransmitters hebben een lager moleculair gewicht dan peptiden en worden aangemaakt vanuit eenvoudige substanties of recht streeks uit het voedsel gehaald.
|
|
|
Klassieke neurotransmitters werken met hoge concentraties en hebben een kortstondig effect.
De neuropeptiden daarentegen bestaan uit |
twee of meer aminozuren die worden aangemaakt door ribosomen in het cellichaam volgens instrncties van chromosomen uit de nucleus.
Ze zijn werkzaam in lage concentraties en hebben een langdurig effect. |
|
|
Algemeen werd aangenomen dat elk neuron over een eigen karakteristieke neu rotransmi tter beschikt. Deze regel staat bekend als |
de wet van Dale (Sir Henri Dale, Nobelprijs in 1936 samen met Otto Loewi, ontdekker van de neurotransmitters acetylcholine en epinephrine) en stelt dat elk neuron gebruik maakt van dezelfde neurotransmitter in al zijn synapsen met andere neuronen.
Natuurlijk kan een neuron wel informatie ontvangen van andere neuronen die andere neurotramitter gebruiken. Recent worden er uitzonderingen op de wet van Dale vastge teld. Vooral neuroactieve peptiden kunnen samen met de klassieke neurotransmitters in de eindplaatje voorkomen.
Het blijft vooralsnog onduidelijk wat de precieze rol van de peptiden onder deze omstandigheden is, maar het wijst alleszins op de enorme complexiteit van de neurobiochemische mechanismen van het neuron. |
|
|
Neurotransmitters oefenen een effect uit op de receptoren van het postsynaptisch membraan.
Receptoren zijn proteïnen die zich met de neurotransmitter kunnen binden . Herinner u dat het effect van een transmitter op een neuron afhankelijk is van het type receptor waarmee de transmitter zich bindt.
Ander gezegd, |
de structuur van bet receptor-effectorcomplex bepaalt het effect van de neurotransmitter.
Veel neurotransmitters binden zich met verschillende type van receptorcomplexen. Zo kunnen dopamine en serotonine zowel excitatorisch als inhi bitori ch zijn afhankelijk van het receptortype waarmee ze zich binden.
Op die manier kunnen neuronen een excitatori cheffect hebben in bepaalde collateralen en een inhibitorisch effect in andere collateralen. |
|
|
Het is mogelijk om de distributie van bepaalde neurotransmitters in de hersenen te bepalen. Sommige neurotransmitters kunnen worden gellokaliseerdin neuronen waarvan de cellichamen in een beperkt aantal kernen zijn gelegen.
Zo wordt bijvoorbeeld norepinephrine in het brein voornamelijk aangemaakt in |
de locus coeruleus .
De locus coeruleus is een cluster van neuronen gesitueerd in de pons van de hersenstam en vormt een deel van de formatio reticularis die betrokken is bij de activatie van het zenuwstelsel.
|
|
|
Het norepinepbrinesysteem reageert binnen enkele seconden op stresserende stimuli en speelt een integratieve rol voor de initiatie van |
biologische en gedragsmatige responsen en dus uiteindelijk voor overleving.
Vanuit de locus coeruleus wordt norepinephrine over grote delen van het brein verspreid waaronder het cerebellum, de hersenstam en de cortex. |
|
|
Drugs met een excitatorisch effect op het centrale zenuwstelsel zoals cocaïne en amfetamine kunnen op twee verschillende manieren dit norepinephrinesysteem beïnvloeden :
|
-ofwel door het effect van norepinephrine ter hoogte van de receptoren te verlengen -ofwel door zich voor te doen als een subtituut voor norepinephrine in de synaptische spleet en alzo het brein te verleiden om zich te gedragen alsof het extreem opgewonden is.
|
|
|
Norepinephrine , epinephrine en dopamine
worden samen .... genoemd |
catecholamines genoemd en zijn biochemisch sterk met elkaar verwant.
Het effect veroorzaakt door epinephrine (vroeger adrenaline genoemd) noemt men een adrenerg effect. Epinephrine is vooral gekend al een sympathische neurotransmitter en zijn rol voor het centraal zenuwstelsel is minder duidelijk.
|
|
|
Het effect veroorzaakt door epinephrine (vroeger adrenaline genoemd) noemt men |
een adrenerg effect. Epinephrine is vooral gekend al een sympathische neurotransmitter en zijn rol voor het centraal zenuwstelsel is minder duidelijk.
|
|
|
De twee belangrijkste dopaminerge baansystemen zijn
-het nigrostriatale en -het mesolimbische systeem.
Het nigrostriatale systeem ontspringt in |
de substantia nigra in de basale ganglia en vormt uitlopers tot in de frontale cortex.
Het mesolimbische systeem ontspringt in het tegmentum en verspreidt zich vooral naar de limbische gebieden van de hersenen.
Dopamine heeft belangrijke invloeden op de motoriek en een tekort van deze neurotransmitter veroorzaakt de ziekte van Parkinson.
Een teveel aan dopamine wordt dan weer geassocieerd met schizofrenie. Een veelvoorkomende bijwerking van (anti)dopa rninerge behandelingen bij schizofrenie is het ontstaan van motorische bijwerkingen die zeer sterk lijken op de motorische disfu ncties van de ziekte van Parkinson. Dopamine is ook betrokken bij de regulatie van emotie via het mesolimbisch systeem. |
|
|
Dopamine heeft belangrijke invloeden op de motoriek en een tekort van deze neurotransmitter veroorzaakt de |
ziekte van Parkinson.
|
|
|
Een teveel aan dopamine wordt dan weer geassocieerd met |
schizofrenie.
Een veelvoorkomende bijwerking van (anti)dopaminerge behandelingen bij schizofrenie is het ontstaan van motorische bijwerkingen die zeer sterk lijken op de motorische disfuncties van de ziekte van Parkinson.
Dopamine is ook betrokken bij de regulatie van emotie via het mesolimbisch systeem. |
|
|
De neurotransmitter acetylcholine wordt vooral aangetroffen in het perifeer zenuwstelsel.
Acetylcholine wordt onder meer gebruikt door |
de nervusvagus voor de parasympathische beïn vloeding van de interne organen waaronder het hart.
Acetylcholine is ook de transmitter die instaat voor de controle over de willekeurige skeletspieren van het lichaam.
|
|
|
Er bestaan twee types van cholinerge receptoren die verschillen in hun agonisten.
-In het ene cholinerge receptortype bootst nicotine de actie van acetylcholine na. -In het andere type is de agonist muscarine (een schimmelderivaat) .
Cholinerge neuronen kunnen derhalve worden beschouwd als |
nicotinisch of muscarinisch en hebben een geheel verschillende werking en eigenschappen.
De nicotinische functie van acetylcholine dient als neurotransmitter ter hoogte van het contact tussen zenuwen en spieren. De muscarinische functie van acetylcholine dient als neuromod ulator in de parasympathische synapsen van het autonoom zenuwstelsel.
Ook in het centraal zenuwstelsel is acetylcholine belangrijk, met name voor geheugen - en intellectuele functies.
Stoornissen bij de regulatie van acetylcholine zijn typisch betrokken bij de ernstige dementiële involutie van de ziekte van Alzheimer. |
|
|
Serotonine en serotonerge neuronen zijn gelokaliseerd in de pons , meer bepaald in de raphe kerncluster.
Serotonine speelt een belangrijke rol in de slaap en afwijkingen in
|
het serotonine ysteem zijn geassocieerd met depressie.
|
|
|
Gaba-aminoboterzuur tenslotte is een belangrijke inhibitorische aminozuurtransmitter en komt in grote hoeveelheden in het hele brein voor. Gaba-aminoboterzuur is vooral betrokken bij |
dopaminerge actie, met inhiberende invloed in zowel nigrostriatale als mesolimbische dopaminerge baansystemen. |